








-
Garlic (Allium spp.), especially the bulb, is commonly consumed and valued for both culinary and medicinal purposes due to its nutritional richness and numerous beneficial bioactive compounds essential for human health[1−3]. Elephant garlic [Allium ampeloprasum var. ampeloprasum (Borrer) Syme], hardneck garlic [A. sativum var. ophioscorodon (Link) Döll], and softneck garlic (A. sativum var. sativum L.) are popular varieties that have been cultivated worldwide[4,5]. In 2022, global garlic production reached 2.91 million tons, valued at 3.43 billion USD. China was the largest producer, contributing 2.13 million tons (73% of world production), followed by India with 0.3 million tons, Bangladesh with 0.05 million tons, and Egypt with 0.03 million tons[6]. Myanmar is the top garlic producer in Southeast Asia followed by Thailand and Indonesia[6]. At every stage of growth, harvesting, and post-harvest storage, garlic is susceptible to various diseases caused by bacteria, fungi, and viruses[7−9]. Diseases can significantly damage garlic bulb production and quality[9,10]. Blue mold disease, caused by Penicillium species, is a common issue affecting garlic bulbs during both the cultivation process and postharvest storage[9,11−14]. This disease can lead to significant customer dissatisfaction and economic losses in garlic production worldwide[9,11,14,15].
In Thailand, the northern part is the main region for garlic cultivation[16]. Nowadays, elephant garlic is a significant vegetable crop extensively cultivated in Thailand. Thus, the area of plantations used for growing garlic has significantly increased in Thailand. However, the incidence and severity of diseases have also increased when plants are grown in sub-optimal areas and unsuitable storage conditions. In 2023, blue mold disease caused by fungi was observed on elephant garlic bulbs during the storage period in Chiang Mai Province in Thailand, with a degree of incidence within the range of 20% to 30%. Importantly, there had been no prior reports of blue mold disease on elephant garlic bulbs in Thailand. Therefore, the objective of this study was to isolate the causal fungal agents of this disease. The isolated fungi were identified using both morphological and molecular data. Pathogenicity tests were conducted, and Koch's postulates were applied to assess the effects of the isolated fungi on asymptomatic elephant garlic bulbs. Moreover, the sensitivity of the isolated fungi to several commercial fungicides was investigated using solid culture techniques.
-
Blue mold disease was observed on elephant garlic bulbs (A. ampeloprasum var. ampeloprasum) throughout the postharvest storage at 25 to 30 °C and 65% to 75% relative humidity over a period of 7 to 14 d in Chiang Mai Province, northern Thailand in 2023 (March to April). Garlic bulbs exhibiting typical symptoms were collected from postharvest storage stores and shipped to the laboratory within 24 h. After being transferred to the laboratory, symptomatic bulbs were examined using a stereo microscope (Nikon H55OS, Tokyo, Japan) and stored in a plastic container with moist filter paper to promote fungal sporulation.
Fungal isolation and morphological study
-
Samples of bulb disease were processed to isolate the fungal causal agents. The single conidial isolation method described by Choi et al.[17] was used to isolate the causal fungi from the lesions. This process was conducted on 1.0% water agar containing 0.5 mg/L streptomycin. The individual germinated conidia were observed after incubation at 25 °C for 24–48 h and then transferred directly onto potato dextrose agar (PDA; CONDA, Madrid, Spain) supplemented with 0.5 mg/L streptomycin under a stereo microscope. Pure cultures were deposited in the Culture Collection of Sustainable Development of Biological Resources (SDBR) Laboratory, Faculty of Science, Chiang Mai University, Thailand. The characteristics of the fungal colonies, including colony morphology, pigmentation, and odor, were examined on PDA, Czapek yeast extract agar (CYA), and malt extract agar (MEA; Difco, France) after incubation in the dark for 7 d at 25 °C. Micromorphological characteristics were assessed using a light microscope (Nikon Eclipse Ni-U, Tokyo, Japan). The Tarosoft® Image Frame Work software was used to measure at least 50 samples for each anatomical structure (such as conidiophores, phialides, and conidia).
DNA extraction, PCR amplification, and sequencing
-
Genomic DNA was extracted from the fungal cultures of each isolate that grew on PDA at 25 °C for 5 d, using a Fungal DNA Extraction Kit (FAVORGEN, Ping-Tung, Taiwan) according to the manufacturer's protocol. Amplification of the internal transcribed spacer (ITS) of ribosomal DNA, β-tubulin (BenA), calmodulin (CaM), and RNA polymerase II second largest subunit (rpb2) genes using ITS5/ITS4[18], Bt2a/Bt2b[19], CF1/CF4[20], and RPB2-5F/RPB2-7CR[21], respectively. The PCR for these four genes was conducted in separate PCR reactions and consisted of an initial denaturation at 95 °C for 3 min, followed by 35 cycles of denaturation at 95 °C for 30 s, annealing at 52 °C for 30 s (ITS and BenA); 51 °C for 1 min (CaM) and 52 °C for 1 min (rpb2), extension at 72 °C for 1 min, and final extension at 72 °C for 10 min on a peqSTAR thermal cycler (PEQLAB Ltd., Fareham, UK). PCR products were checked on 1% agarose gel electrophoresis and purified using a PCR clean-up Gel Extraction NucleoSpin® Gel and a PCR Clean-up Kit (Macherey-Nagel, Düren, Germany), according to the manufacturer's instructions. The purified PCR products were directly sequenced. Sequencing reactions were performed, and the above-mentioned PCR primers were employed to automatically determine the sequences in the Genetic Analyzer at the 1st Base Company (Kembangan, Malaysia).
Sequence alignment and phylogenetic analyses
-
The analysis of the ITS, BenA, CaM, and rpb2 sequences was conducted with the use of similarity searches employing the BLAST program available at NCBI (
http://blast.ddbj.nig.ac.jp/top-e.html , accessed on 10 July 2024). The sequences from this study and those obtained from previous studies, together with sequences downloaded from the nucleotide GenBank database are listed in Table 1. Multiple sequence alignment was performed with MUSCLE[22] and improved where necessary using BioEdit v. 6.0.7[23]. Finally, the combination datasets of ITS, BenA, CaM, and rpb2 sequences were performed.Table 1. Details of sequences in Penicillium section Fasciculata used in molecular phylogenetic analysis.
Penicillium species Strain/isolate GenBank accession number ITS BenA CaM rpb2 P. albocoremium CBS 472.84T AJ004819 AY674326 KUJ896819 KU904344 P. allii CBS 131.89T ‒ AY674331 KU896820 KU904345 P. allii SDBR-CMU499 PP998350 PQ032853 PQ032856 PQ032859 P. allii SDBR-CMU500 PP998351 PQ032854 PQ032857 PQ032860 P. allii SDBR-CMU501 PP998352 PQ032855 PQ032858 PQ032861 P. aurantiogriseum CBS 324.89 AF033476 AY674296 KU896822 JN406573 P. camemberti MUCL 29790T AB479314 FJ930956 KU896825 JN121484 P. cavernicola CBS 100540T MH862709 KJ834439 KU896827 KU904348 P. caseifulvum CBS 101134T MH862722 AY674372 KU896826 KU904347 P. commune CBS 311.48T AY213672 AY674366 KU896829 KU904350 P. concentricum CBS 477.75T KC411763 AY674413 DQ911131 KT900575 P. coprobium CBS 561.90T DQ339559 AY674425 KU896830 KT900576 P. discolor CBS 474.84T OW986149 AY674348 KU896834 KU904351 P. echinulatum CBS 317.48T MH856364 AY674341 DQ911133 KU904352 P. freii CBS 476.84T MH861769 KU896813 KU896836 KU904353 P. gladioli CBS 332.48T AF033480 AY674287 KU896837 JN406567 P. glandicola CBS 498.75T AB479308 AY674415 KU896838 KU904354 P. griseofulvum CBS 185.27T AF033468 AY674432 JX996966 JN121449 P. hirsutum CBS 135.41T AY373918 AF003243 KU896840 JN406629 P. hordei CBS 701.68T MN431391 AY674347 KU896841 KU904355 P. italicum CBS 339.48T KJ834509 AY674398 DQ911135 ‒ P. melanoconidium CV1331 JX091410 JX091545 JX141587 KU904358 P. neoechinulatum CBS 101135T JN942722 AF003237 KU896844 JN985406 P. nordicum DTO 098-F7 KJ834513 KJ834476 KU896845 KU904359 P. palitans CBS 107.11T KJ834514 KJ834480 KU896847 KU904360 P. polonicum CBS 222.28T AF033475 AY674305 KU896848 JN406609 P. solitum CBS 424.89T AY373932 AY674354 KU896851 KU904363 P. thymicola CBS 111225T KJ834518 AY674321 FJ530990 KU904364 P. tricolor CBS 635.93T MH862450 AY674313 KU896852 JN985422 P. ulaiense CBS 210.92T KC411695 AY674408 KUB96854 KU904365 P. verrucosum CBS 603.74T AY373938 AY674323 DQ911138 JN121539 P. vulpinum CBS 126.23T AF506012 KJ834501 KU896857 KU904367 Ex-type species are indicated by the superscript letters as 'T'. '−' indicates the absence of sequencing information in GenBank. The fungal isolates and sequences obtained in this study are in bold. For phylogenetic analyses, Penicillium italicum (CBS 339.48) and P. ulaiense (CBS 210.92) were selected as the outgroup. The maximum likelihood (ML) analysis was carried out using RAxML-HPC2 version 8.2.12 on the GTRCAT model with 25 categories and 1000 bootstrap (BS) replications[24,25] via the CIPRES web portal. Bayesian inference (BI) analysis was performed with MrBayes v. 3.2.6 software for Windows[26]. The best substitution model for BI analysis was estimated using the jModelTest 2.1.10[27] by employing the Akaike information criterion (AIC). Bayesian posterior probability (PP) was determined by Markov Chain Monte Carlo Sampling (MCMC). Four simultaneous Markov chains were run for a million generations with random initial trees, wherein every 100 generations were sampled. The first 25% of generated trees representing the burn-in phase of the analysis were eliminated, while the remaining trees were used for calculating PP in the majority-rule consensus tree. The phylogenetic trees were visualized using FigTree v1.4.0[28].
Pathogenicity tests
-
Conidia from fungal isolates cultivated for two weeks on PDA were used in this experiment. Healthy commercial elephant garlic bulbs were washed thoroughly, and then their surfaces were sterilized by soaking them for 5 min in a sterile sodium hypochlorite solution with a concentration of 1.5% (v/v). Following that, sterile distilled water was used to wash them three times. The bulbs were allowed to air-dry at room temperature (25 ± 2 °C) for 10 min after surface disinfection. Following the air-drying process, a quantity of 10 μL of a conidial suspension (1 × 106 conidia/mL) in sterile water from each fungal isolate was dropped onto each bulb. Consequently, sterile distilled water was used as an inoculant for the control. The inoculated bulbs were placed in individual 4 L sterile plastic boxes maintained at 80% relative humidity. These containers were kept in a growth chamber at a temperature of 25 °C under a 12-h light cycle for one week. A total of ten replicates were used for each treatment, which was repeated twice under the same conditions. The disease symptoms were observed. Confirmation of Koch's postulates was achieved by re-isolating the fungi through the isolation method from any lesions that occurred on the inoculated bulbs.
Screening of commercial fungicides against the causal agent
-
Seven commercially available fungicides, including benalaxyl-M (4%) + mancozeb (65%) (Fantic M WG®, Thailand), captan (Captan 50®, Thailand), carbendazim (Dazine®, Thailand), copper oxychloride (Copina 85 WP®, Thailand), difenoconazole (12.5%) + azoxystrobin (20%) (Ortiva®, Thailand), difenoconazole (Score®, Thailand), and mancozeb (Newthane M-80®, Thailand), were examined in this study according to the approach indicated through previous studies[29,30]. The fungicides used in this study were available commercially in Thailand and were approved for usage. The in vitro applications of benalaxyl-M + mancozeb, captan, carbendazim, copper oxychloride, difenoconazole + azoxystrobin, difenoconazole, and mancozeb were recommended at dosages of 1,380, 750, 750, 1,700, 243.75, 187.5, and 1,200 ppm, respectively, according to the labels for each fungicide. The final concentration was obtained by preparing each fungicide and adding it to an autoclaved PDA. Each fungicide was used in three different dosages: half-recommended, recommended, and double-recommended. The test media were inoculated with mycelial plugs (5 mm in diameter) that had been cultivated on PDA in the dark at 25 °C for one week. The control did not add any fungicide. The plates were maintained in darkness at a temperature of 25 °C. Following one week of incubation, the mycelial growth of each isolate was evaluated on individual plates and a comparison was made between the growth in PDA supplemented with fungicides and the growth observed in the control. The calculation of the percentage growth inhibition for each isolate was performed using the formula provided by Pandey et al.[31]. Each isolate was classified as sensitive (> 50% inhibition), insensitive (< 50% inhibition), or totally inhibited (100% inhibition) based on their growth inhibition rates[30,31]. Five replicates were conducted for each fungicide and fungal isolate, and the experiments were independently repeated twice under the same biological conditions.
Statistical analysis
-
The Shapiro-Wilk test in SPSS software version 26 was used to examine data from the two repeated fungicide sensitivity experiments at a significant level of p < 0.05 to perform the normality test. The results indicated non-significant findings, so the data from these repeated experiments were assessed for the assumptions of one-way analysis of variance (ANOVA). Duncan's Multiple Range Test (DMRT) was then used to identify significant differences at p ≤ 0.05.
-
Initial symptoms, water-soaked areas on the outer surface of scales were observed. Later, white mycelium and blue powdery mold develop on the surface of the lesions (Fig. 1a). These lesions appear as brown, tan, or grey colored areas when the bulbs are cut. In advanced stages, infected bulbs disintegrated into a watery rot.
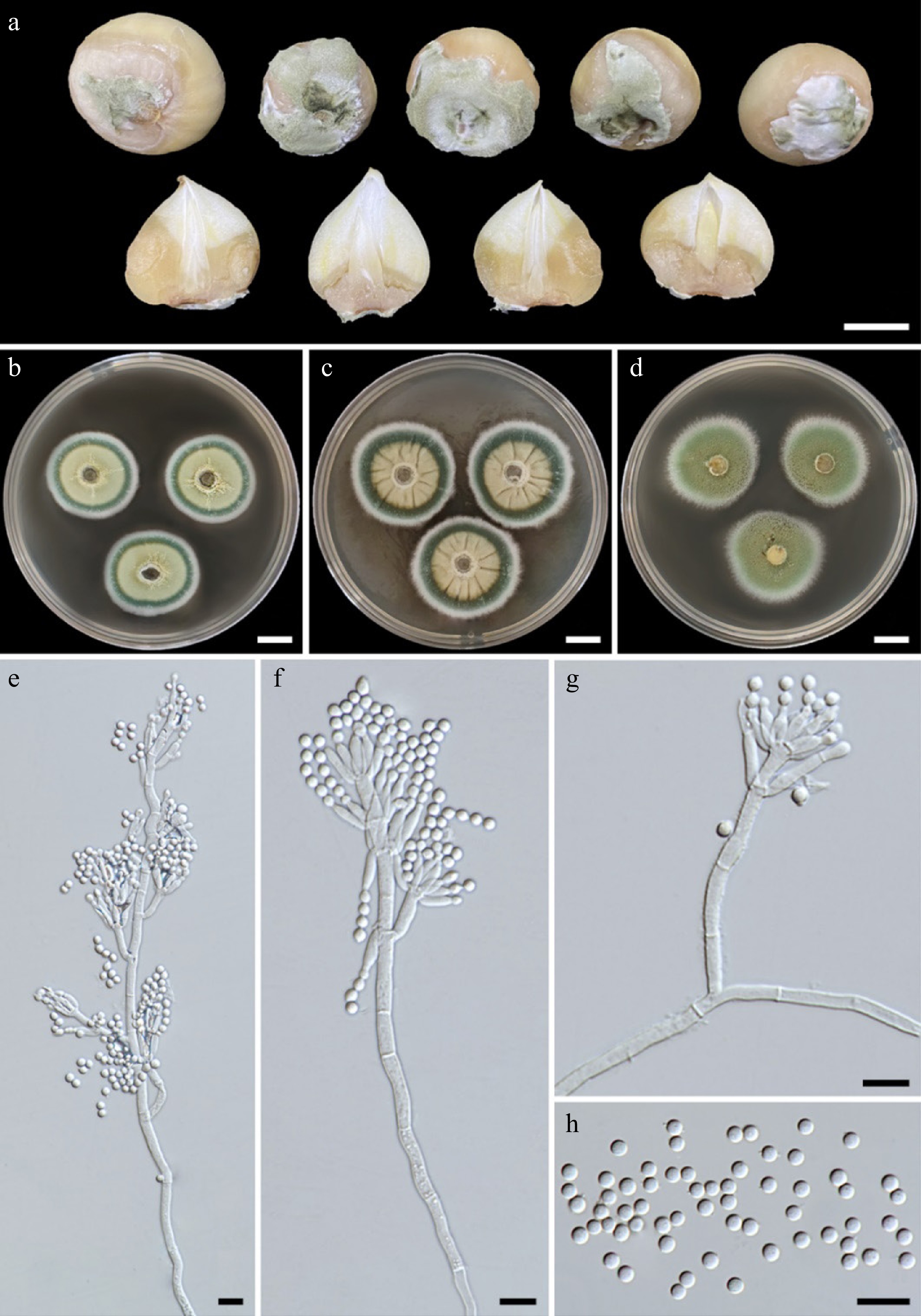
Figure 1.
(a) Natural symptoms of blue mold disease on bulbs of elephant garlic by Penicillium allii . Colonies of Penicillium allii SDBR-CMU499 after incubation at 25 °C for one week. (b) PDA. (c) CYA. (d) MEA. (e)–(g) Conidiophores. (h) Conidia. Scale bars: (a)–(d) = 10 mm, (e)–(h) = 10 μm.
Fungal isolation and morphological characteristics
-
Three fungal isolates (CMU499, CMU500, and CMU501) with similar morphology were obtained and deposited at the SDBR-CMU under accession numbers SDBR-CMU499, SDBR-CMU500, and SDBR-CMU501, respectively. Colonies PDA, CYA, and MEA were 29–32, 32–37, and 33–37 mm in diameter, respectively after incubation for one week at 25 °C (Fig. 1b–d). Colonies on PDA and MEA were white, flat with entire edges, conidium turquoise, white in the center, dull green at the margins; reverse pale yellow for PDA and yellow-brown for MEA. Colonies on CYA were white, flat with entire edges, conidium dull green; reverse white. All fungal isolates could produce conidiophores, and phialides, and sporulate in all of the agar media. Conidiophores terverticillate (Fig. 1e–g). Stipes rough-walled, 13.2–181.2 × 2.3–3.9 μm. Rami one or two, rough-walled and appressed or divergent, 8.4–24.7 × 2.5–4.6 μm. Metulae divergent, in verticils of 2–4, 8–19.1 × 2.3–4.6 μm. Phialides ampulliform, in verticils of 3 to 9, 6–17.9 × 1.7–6.9 μm. Conidia globose, 2.6–4.3 μm in diameter, smooth-walled, dull green (Fig. 1h). Based on these morphological characteristics, all fungal isolates were initially identified as belonging to Penicillium[32−35]. Fungal identification was then further confirmed using multi-gene phylogenetic analyses.
Phylogenetic analysis
-
The ITS, BenA, CaM, and rpb2 sequences obtained from three fungal isolates in this study have been deposited in GenBank (Table 1). According to the BLAST results, all fungal isolates were identified as members of the Penicillium section Fasciculata. The combined ITS, BenA, CaM, and rpb2 sequences dataset consists of 32 taxa, and the aligned dataset includes 2,399 characters comprising gaps (ITS: 1–553, BenA: 554–927, CaM: 928–1,442, and rpb2: 1,443–2,399). The best-scoring RAxML tree was established with a final ML optimization likelihood value of –9,279.455311. Accordingly, the matrix contained 612 distinct alignment patterns with 5.04% undetermined characters or gaps. The estimated base frequencies were found to be: A = 0.235164, C = 0.269054, G = 0.262373, and T = 0.233408; substitution rates AC = 1.298254, AG = 4.466196, AT = 1.340638, CG = 0.787838, CT = 9.362594, and GT = 1.00000. The values of the gamma distribution shape parameter alpha and the Tree-Length were 0.580385 and 0.524602, respectively. Additionally, BI analysis yielded a final average standard deviation of 0.002257 for the split frequencies at the end of all MCMC generations. Regarding topology, the phylogenetic trees generated from both ML and BI analyses were similar. Consequently, the phylogenetic tree obtained from the ML analysis was selected and is displayed in Fig. 2. The results indicated that three fungal isolates SDBR-CMU499, SDBR-CMU500, and SDBR-CMU501 clustered with P. allii CBS 131.89 (ex-type strain) with strong statistical (100% BS and 1.0 PP) supports. Therefore, all fungal isolates obtained in this were identified as P. allii based on morphological and molecular data.
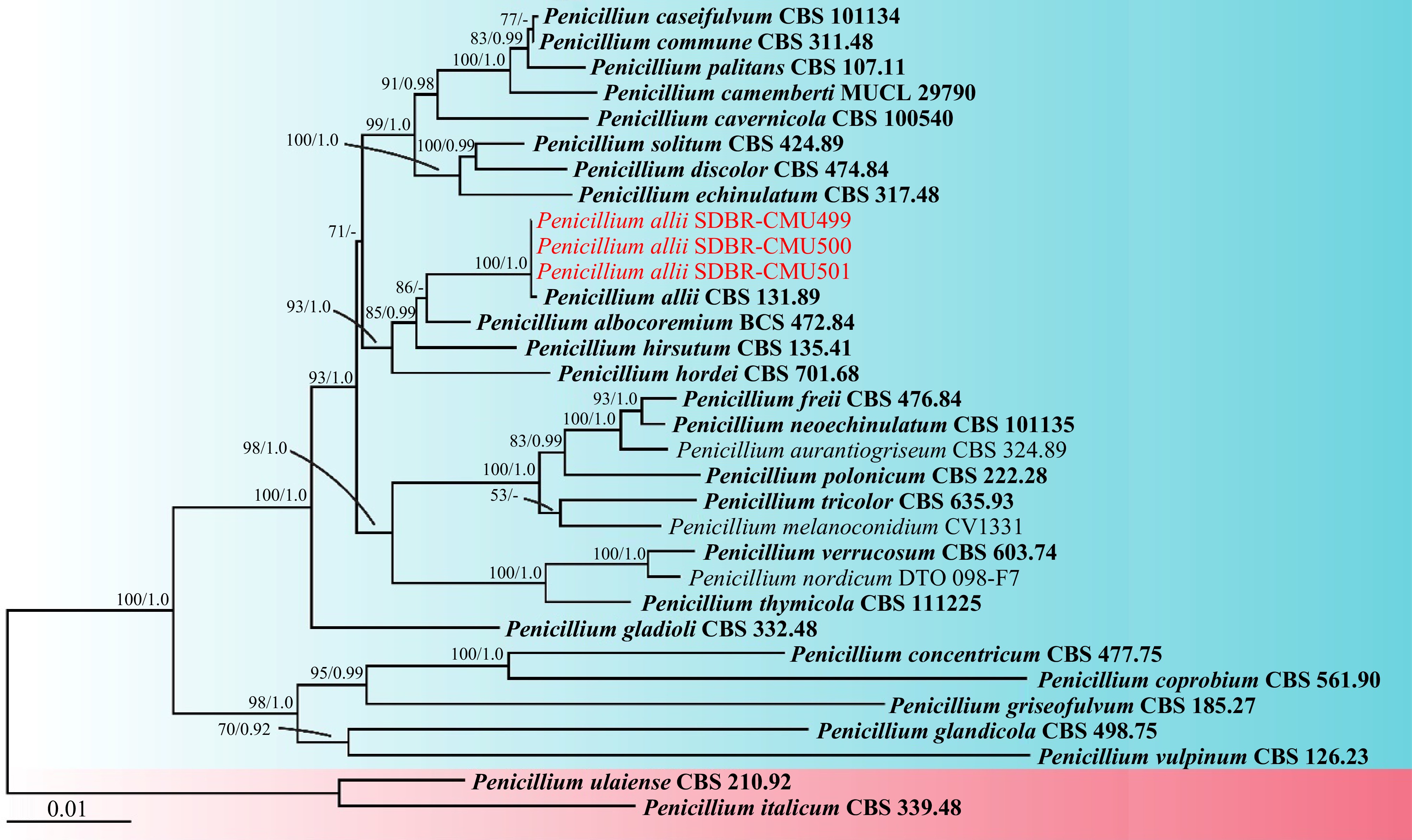
Figure 2.
Phylogram derived from maximum likelihood analysis of the combined ITS, BenA, CaM, and rpb2 sequences of 30 taxa in the Penicillium section Fasciculata and two taxa in the Penicillium section Penicillium. Penicillium italicum CBS 339.48 and P. ulaiense CBS 210.92 were used as outgroups. Bootstrap values ≥ 50% (left) and Bayesian posterior probabilities ≥ 0.90 (right) are displayed above nodes. The scale bar represents the expected number of nucleotide substitutions per site. The sequences of fungal species obtained in this study are in red. The ex-type strain are in bold.
Pathogenicity tests
-
The initial symptoms appeared on bulbs of elephant garlic 3 d after being inoculated. After 7 d, all inoculated bulbs displayed powdery mold at their centers, surrounded by orange-brown water-soaked lesions (Fig. 3b‒d). Whereas, control bulbs were asymptomatic (Fig. 3a). Penicillium allii was consistently reisolated from the inoculated bulbs on PDA to complete Koch's postulates.
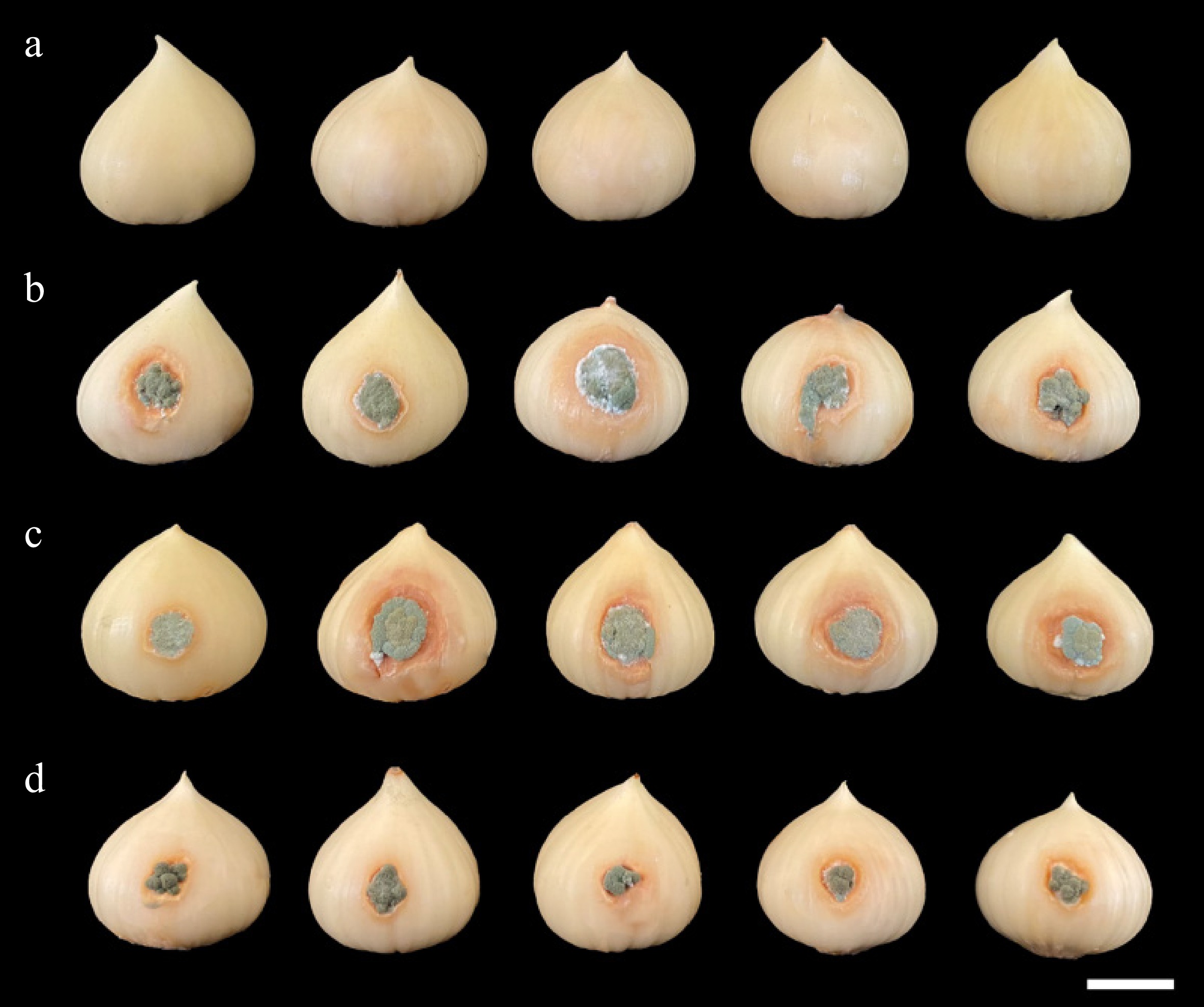
Figure 3.
Pathogenicity test using Penicillium allii SDBR-CMU499, SDBR-CMU500, and SDBR-CMU501 on bulbs of elephant garlic after one week inoculation at 25 °C. (a) Control bulbs treated with sterile distilled water instead of inoculum. Blue mold disease on bulbs of elephant garlic after inoculation of isolate (b) SDBR-CMU499, (c) SDBR-CMU500, and (d) SDBR-CMU501. Scale bar: 10 mm.
Reactions of commercial fungicides against Penicillium allii
-
Seven commercially available fungicides in Thailand were tested in this study. After one week, the mycelial growths in response to the fungicides at three different dosages, including half-recommended (1/2RD), recommended (RD), and double-recommended (2RD) were calculated and presented in Table 2. The results revealed that the growth inhibition values varied across different fungicides, dosages, and fungal isolates. Data on the percentage of mycelial inhibition for each fungal isolate, related to the fungicides, passed the normality test (Shapiro-Wilk test, p < 0.001), thereby assuming normal distributions. Therefore, ANOVA followed by DMRT (p ≤ 0.05) was used to identify significant differences. The findings indicated that all fungal isolates were completely inhibited by carbendazim, difenoconazole + azoxystrobin, and difenoconazole at all tested dosages (Table 2). In the tests for captan and mancozeb, all isolates demonstrated sensitivity to 2RD. Therefore, based on the recommended dosages, carbendazim, difenoconazole + azoxystrobin, and difenoconazole could be effectively applied to control this pathogen.
Table 2. Percentage of mycelial inhibition and reactions of three isolates of Penicillium allii against fungicides.
Fungicides Dosages Inhibition of mycelial growth (%)* Reaction SDBR-CMU499 SDBR-CMU500 SDBR-CMU501 Benalaxyl-M + mancozeb 1/2RD 30.08 ± 1.41 c 29.27 ± 2.25 c 30.08 ± 2.53 c Insensitive RD 55.28 ± 1.41 b 56.10 ± 3.62 b 60.16 ± 1.67 b Sensitive 2RD 82.11 ± 2.82 a 83.74 ± 1.41 a 83.74 ± 3.45 a Sensitive Captan 1/2RD 2.44 ± 3.50 c 1.63 ± 2.41 c 2.44 ± 1.25 c Insensitive RD 4.88 ± 2.25 b 4.88 ± 2.23 b 5.69 ± 2.30 b Insensitive 2RD 72.36 ± 1.60 a 73.93 ± 3.45 a 73.98 ± 2.82 a Sensitive Carbendazim 1/2RD 100 ± 0 a 100 ± 0 a 100 ± 0 a Inhibition RD 100 ± 0 a 100 ± 0 a 100 ± 0 a Inhibition 2RD 100 ± 0 a 100 ± 0 a 100 ± 0 a Inhibition Copper oxychloride 1/2RD 61.79 ± 1.60 c 58.54 ± 1.05 c 56.91 ± 1.41 f Sensitive RD 68.29 ± 1.20 b 68.29 ± 2.05 b 68.29 ± 2.54 d Sensitive 2RD 100 ± 0 a 100 ± 0 a 100 ± 0 a Inhibition Difenoconazole + azoxystrobin 1/2RD 100 ± 0 a 100 ± 0 a 100 ± 0 a Inhibition RD 100 ± 0 a 100 ± 0 a 100 ± 0 a Inhibition 2RD 100 ± 0 a 100 ± 0 a 100 ± 0 a Inhibition Difenoconazole 1/2RD 100 ± 0 a 100 ± 0 a 100 ± 0 a Inhibition RD 100 ± 0 a 100 ± 0 a 100 ± 0 a Inhibition 2RD 100 ± 0 a 100 ± 0 a 100 ± 0 a Inhibition Mancozeb 1/2RD 19.51 ± 2.44 c 22.76 ± 3.73 c 21.14 ± 1.41 c Insensitive RD 47.15 ± 1.45 b 47.15 ± 2.82 b 43.90 ± 2.44 b Insensitive 2RD 58.54 ± 2.44 a 54.47 ± 1.42 a 55.28 ± 1.45 a Sensitive * Results are means of five replicates ± standard deviation with the independently repeated twice. Data with different letters within the same column for each fungal isolate and fungicide indicate a significant difference at p ≤ 0.05 according to Duncan's multiple range test. 1/2RD, RD, and 2RD indicate half of the recommended dosage, recommended dosage, and double the recommended dosage, respectively. -
Penicillium species are widely recognized as one of the most significant genera, known to cause major diseases in numerous economically valuable crops cultivated worldwide, including garlic[9,10,12,13,36]. Traditionally, Penicillium species have been identified using both macromorphological and micromorphological characteristics. However, morphological traits alone are insufficient to differentiate closely related Penicillium species due to the extensive range of morphological variations. Therefore, molecular techniques are crucial for accurately identifying Penicillium at the species level. Several previous studies have utilized a combination of ribosomal DNA (ITS) and protein-coding genes (BenA, CaM, rpb1, and rpb2) as powerful tools to identify Penicillium species since species-level identification remained unresolved when used solely on the ribosomal DNA gene[32,34,35,37]. In this study, three isolates of P. allii were obtained from the rot lesions of blue mold disease on elephant garlic bulbs in northern Thailand. The identification of this fungal species followed methods similar to those used for identifying Penicillium, which involve combining phylogenetic analysis of multiple genes with their morphological characteristics.
In this study, Koch's postulates were fulfilled by conducting pathogenicity tests on all isolates of P. allii. The findings demonstrate that postharvest blue mold disease on elephant garlic bulbs in northern Thailand, caused by P. allii identified in this study resembles the disease caused by previously identified Penicillium pathogens, particularly P. hirsutum, which affects garlic bulbs worldwide[11−13,36]. Penicillium polonicum has been reported as a causal agent of blue mold on stored garlic bulbs in Pakistan[38]. Penicillium allii was known to cause postharvest blue mold disease on garlic bulbs in Argentina[15,39,40]. In the USA, P. albocoremium, P. expansum, P. glabrum, P. paraherquei, and P. radicicola can cause blue mold on garlic bulbs[13]. In Korea, blue mold disease on garlic bulbs caused by P. hirsutum has been reported[41]. Five Penicillium species, namely P. allii, P. glabrum, P. italicum, P. polonicum, and P. psychrotrophicum were identified and confirmed as postharvest pathogens causing blue mold rot of garlic in Serbia[42]. Recently, P. allii was the most virulent pathogen causing blue mold disease of elephant garlic bulbs in Italy, accounting for 95% of cases, followed by P. citrinum (4%) and P. brevicompactum (1%)[43]. Before this study, there were no reports of blue mold disease on elephant garlic bulbs in Thailand. Thus, this represents the first report of postharvest blue mold disease on elephant garlic bulbs caused by P. allii in Thailand.
To manage and control fungal-caused plant diseases, a variety of fungicides have been used. Several studies have documented the effectiveness of fungicides in affecting sensitive, resistant strains of plant pathogenic fungi, particularly those in the Penicillium species, on their in vitro mycelial growth[44−46]. In this study, the sensitivity and inhibition of P. allii to fungicides varied among different fungicides and dosages. These findings are consistent with previous studies, which reported that the sensitivity and inhibition of Penicillium species to fungicides varies based on the type and dosage of the fungicide, as well as fungal species[46−48]. Before this study, prochloraz had been used against P. allii to control diseases related to sprouting germination in Europe[49]. In this study, carbendazim, difenoconazole + azoxystrobin, and difenoconazole at both half and recommended dosages exhibited complete inhibition of P. allii. The information on the in vitro inhibition, sensitivity, and resistance of fungicides against P. allii, which causes postharvest blue mold disease on elephant garlic bulbs, would be beneficial for in vivo applications and for managing this disease in Thailand and globally. However, environmental factors and the fungicide's metabolism in the plant can cause the results of in vitro fungicide testing to differ from in vivo responses. Therefore, further studies are required to conduct in vivo fungicide sensitivity and disease inhibition assays based on the in vitro findings. Additionally, several previous studies have established that fungicide-resistant strains are a result of both excessive and prolonged fungicide treatment[50−52]. Utilizing biological control agents, rotating crops, adhering to fungicide treatment guidelines, and maintaining cleanliness in fields, equipment, and storage spaces are all essential components of a comprehensive strategy to reduce fungicide resistance in fungi[9,50,53,54].
-
Garlic blue mold disease, caused by Penicillium species, leads to significant economic losses during postharvest storage worldwide. In the present study, P. allii was isolated from infected bulbs of elephant garlic in northern Thailand. The identification of this fungi involved the analysis of their morphological characteristics and conducting multi-gene phylogenetic analyses. The assessment of pathogenicity for P. allii showed similar symptoms throughout the artificial inoculation process, as observed during the postharvest storage period. Therefore, this study represents the first report of elephant garlic blue mold disease caused by P. allii in Thailand. In the fungicide screening test, carbendazim, difenoconazole + azoxystrobin, and difenoconazole were found to effectively control this pathogen at both half and full recommended dosages. Thus, half of the recommended dosages can be used in managing this disease, serving as a guideline for prevention and helping to reduce pathogen resistance to fungicides. The findings of this study will enhance our understanding of postharvest blue mold disease in elephant garlic bulbs and provide insights for developing effective management strategies and prevention methods to minimize significant economic losses. Further research on the epidemiology of this disease would be required for effective monitoring, prevention, and control.
The authors sincerely appreciate the financial support provided by Chiang Mai University, Thailand and the University of Phayao Innovation Fund [Fundamental Fund 2024 (227/2567)], Thailand.
-
The authors confirm contribution to the paper as follows: conceptualization: Suwannarach N, Khuna S; formal analysis: Suwannarach N, Khuna S, Chaiwong K, Senwanna C, Kumla J; investigation, methodology: Suwannarach N, Khuna S, Chaiwong K; resources: Suwannarach N, Khuna S, Chaiwong K; software: Khuna S, Chaiwong K, Senwanna C, Kumla J; validation: Suwannarach N, Khuna S, Senwanna C, Nuangmek W; data curation: Khuna S, Senwanna C, Nuangmek W, Kumla J; visualization: Khuna S, Chaiwong K; writing–original draft: Suwannarach N, Khuna S, Nuangmek W, Kumla J; writing–review & editing: Suwannarach N, Khuna S, Chaiwong K, Senwanna C, Nuangmek W, Kumla J; supervision, project administration: Suwannarach N; funding acquisition: Suwannarach N, Nuangmek W. All authors have read and agreed to the published version of the manuscript.
-
The DNA sequences generated in this study have been submitted to GenBank and can be accessed through the accession numbers provided in the paper.
-
The authors declare that they have no conflict of interest. Nakarin Suwannarach and Jaturong Kumla are the Editorial Board members of Studies in Fungi who are blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer review handled independently of these Editorial Board members and the research groups.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Suwannarach N, Khuna S, Chaiwong K, Senwanna C, Nuangmek W, et al. 2024. Identification and fungicide sensitivity of the blue mold pathogen in postharvest-stored elephant garlic bulbs in Thailand. Studies in Fungi 9: e015 doi: 10.48130/sif-0024-0015 

Identification and fungicide sensitivity of the blue mold pathogen in postharvest-stored elephant garlic bulbs in Thailand
- Received: 05 November 2024
- Revised: 25 November 2024
- Accepted: 26 November 2024
- Published online: 20 December 2024
Abstract: Blue mold disease is one of the most important postharvest diseases affecting garlic bulbs. In 2023, this disease was found on bulbs of elephant garlic [Allium ampeloprasum var. ampeloprasum (Borrer) Syme] in Chiang Mai Province, Thailand, during the postharvest storage period. Three fungal isolates were obtained and identified as Penicillium allii based on morphological characteristics and phylogenetic analysis of combined sequences of the internal transcribed spacer (ITS) of ribosomal DNA, β-tubulin (BenA), calmodulin (CaM), and RNA polymerase II second largest subunit (rpb2) genes. In the pathogenicity test, garlic bulbs inoculated with the isolated fungi exhibited symptoms similar to those observed during the postharvest storage period. In the fungicide screening test, carbendazim, difenoconazole + azoxystrobin, and difenoconazole effectively completely inhibited this fungus at both half and recommended dosages, while the fungus showed insensitivity to captan and mancozeb. Additionally, double-recommended dosages of carbendazim, copper oxychloride, difenoconazole combined with azoxystrobin, and difenoconazole alone exhibited complete inhibition of the fungus. To the best of our knowledge, this is the first report of postharvest blue mold disease on elephant garlic bulbs caused by P. allii in Thailand. Furthermore, the results of the fungicide sensitivity screening could help in developing effective management strategies for controlling postharvest blue mold disease on elephant garlic bulbs caused by P. allii.
-
Key words:
- Crop protection /
- Fungal pathogen /
- Garlic /
- Pathogen identification /
- Postharvest diseases