









-
Tea, a significant cash crop in China, faces a growing array of diseases due to the expanding cultivation areas. Being a monocultural perennial crop, tea plant thrive in warm, moist regions with consistent rainfall, creating an environment conducive to pathogen proliferation. Currently, over 400 types of pathogens have been identified in tea plant, predominantly fungi and bacteria, with viruses and algae being less prevalent[1]. Given that the leaf is the primary yield of the tea plant, the impact of leaf diseases holds greater importance. It has been observed that diseased leaves exhibit a significant reduction in polyphenols, catechins, and amino acids in comparison to healthy leaves[2]. Tea leaf diseases, including tea plant anthracnose, blister blight, grey blight, and red leaf spot, are prevalent in tea plantations[3]. Among these, anthracnose, the most widespread disease in tea gardens, is attributed to infection by the Deuteromycotina colletotrichum. After infection, leaves exhibit dark green water stains initially, followed by the gradual emergence of brown or reddish-brown spots. Subsequently, the affected areas transition to a grayish-white color. The diseased leaves become fragile and can easily break. Severe outbreaks can lead to extensive damage, directly impacting the quality and yield of spring tea the following year[4].
Tea disease control methods encompass agricultural, chemical, and biological approaches. Agricultural control involves strategic fertilization, precise pruning, and systematic harvesting in tea gardens to manage diseases. Chemical agents remain the primary means of disease control due to their cost-effectiveness, broad applicability, and high efficacy. However, the excessive use of pesticides can lead to the development of resistance in diseases and pests, causing environmental pollution and leaving behind pesticide residues that pose a threat to human health. As living standards improve, there is a growing focus on the quality and safety of tea, the conservation of biodiversity in tea gardens, and the mitigation of environmental pollution. The use of biological control methods has become increasingly crucial for the sustainable growth of the tea industry, drawing attention due to its non-resistance and safety benefits. Notably, the investigation of pathogen antagonism has garnered escalating interest[5]. Lu et al.[6] identified 35 Trichoderma isolates from the rhizosphere soil of a healthy tea garden, with seven exhibiting antagonistic activity against the tea grey blight pathogen. Zhu et al.[7] isolated Bacillus amylolytica JT68 from the tea plant rhizosphere, which demonstrated a 50.73% inhibitory rate of volatile organic compounds against tea plant anthracnose. Moreover, JT68 exhibited inhibitory rates ranging from 70.0% to 93.2% against various plant pathogens including Magnaporthe oryzae, Colletotrichum higginsianum, and Verticillium dahliae. Currently, JT68 is utilized in the production of tea leaf fertilizer. Yang et al.[8] through the organization separation of tea grey blight disease in the separate identification of microorganisms, in addition to screening to pathogen Pseudopestalotiopsis theae, also to screen the inhibitory action to the strains of pathogenic bacteria of kc-6, identified as Bacillus amyloliquefaciens. In addition to inhibiting the growth of pathogenic bacteria, kc-6 also had a better inhibitory effect on Phomopsis vexans, Phytophthora melonis and Fusarium graminearum.
In nature, plants are actually in a symbiotic state with microorganisms. Endophytes refer to the microorganisms residing in different plant organs and tissues, which do not induce disease in the plant temporarily[9]. Endophytes encompass epiphytic fungi, actinomycetes, and bacteria. They are widely distributed and have developed a mutually beneficial symbiotic relationship through long-term co-evolution with plants[10]. Endophytes play a crucial role in protecting plants against pathogenic fungi, bacteria, viruses, and pests, enhancing the host's stress resistance under adverse conditions. For instance, non-pathogenic foliar fungi in Populus trichocarpa have been shown to influence the severity of Melampsora rust disease[11]. Endophytes also can produce secondary metabolites that inhibit pathogen effects on plants, such as Trichoderma activating host defense pathways to mitigate disease impact[12]. Additionally, endophytes can also induce systemic acquired resistance (SAR) or systemic resistance (ISR) against pathogen invasion[13−15]. Some of them secrete low concentrations of metabolites to antagonize pathogens[16].They colonizing almost all plants and conferring higher resistance to colonized hosts compared to non-colonized plants[17].
Current research on tea anthracnose primarily focuses on isolating and identifying pathogens, with limited studies on biological control methods and few antagonists utilized for managing this disease in tea plant. This study isolated and identified epiphytic fungi present in healthy tea leaves, screening for those with potent inhibitory effects. The findings offer a potential approach for biologically controlling tea plant anthracnose and serve as a valuable reference for developing biocontrol agents against this disease in tea plant.
-
One-year-old Camellia sinensis 'Longjing 43' cutting seedlings were acquired from the Tea Sci-tech Demonstration Base of Northwest A&F University in Xixiang, Shaanxi Province, China. The second and third leaves, which were healthy and mature on the new branches of the tea plant with the same leaf size, were selected for the experiment. The potato dextrose agar (PDA) medium was purchased from Beijing Solarbio Science & Technology Co., Ltd. (Beijing, China).
Pathogenic fungi
-
In the field, tea leaves with brown dead, irregular water stains on the edges, and other typical symptoms of anthracnose were collected. Fresh diseased leaves of tea plant with typical diseased spots were selected, and four pieces of leaf tissue (4 mm2) at the junction were cut with sterilized scissors (at the junction of the healthy and diseased regions). The cut diseased leaf tissue was disinfected with 0.1% H2O2 surface for 2 min, then transferred to 75% ethanol for 20 s, and rinsed with sterilized water three consecutive times for 1 min each time. The leaves were placed on sterile paper and excess water was removed from the leaf tissue. The sterilized leaves were transferred to PDA medium in a super-clean workbench, with four pieces in each dish. After labeling, the leaves were placed in a 28 °C incubator for dark culture. After the fungal growth, the mycelium was selected for purification, and then the plant pathogenic fungal were obtained.
DNA was extracted using the CTAB method (200 mM Tris-HCl, pH 8.0, 50 mM EDTA, 20 mM NaCl, and 1% SDS)[18,19] and internal transcribed spacer (ITS) region was amplified by ITS1/4 primers (F: 5'-TCCGTAGGTGAACCTGCGG-3', R: 5'-TCCTCCGCTTATTGATATGC-3'). The procedures used for the amplification of ITS sequences were as follows: predenaturation at 95 °C for 3 min, denaturation at 95 °C for 30 s, annealing at 52 °C for 45 s, extension at 72 °C for 90 s, extension at 72 °C for 10 min after 35 cycles. The size of the amplified fragment was detected by 1.5 μL PCR product in 1.5% agarose gel electrophoresis, which was purified and sequenced in a sangon (Shanghai, China). The sequences were blast in NCBI GenBank and were identified as Colletotrichum camelliae with 99% similarity.
The fully unfolded 'Longjing 43' second and third leaves were selected with 75% ethanol for surface disinfection, scrubbed with sterile water and air dried naturally. The sterilized head was used to damage the leaves, avoiding the vein position. A hole punch with a diameter of 6 mm was used to punch holes at the edge of fresh mycelia. The mycelia pieces were inoculated on the damaged part of the leaves, and fresh PDA medium was used as the control. Each strain had at least six duplicates. The inoculated leaves were wrapped in plastic wrap and cultured in an artificial climate chamber (25 ± 2 °C; humidity above 70%). To verify whether the results satisfied Koch's rule, the infected leaves were isolated and re-identified to determine whether they were pre-inoculation strains[20].
Purification and morphological observation of antagonistic fungus
-
Fungus with obvious antagonistic effects against pathogens on PDA medium were selected in the process of successive transfer culture. Single hyphal tips were transferred to a new PDA and cultured for 5 d at 25 °C in the dark. The size and morphology of fungal spores, hyphae, and other reproduction organs were observed with a microscope (Olympus BX51, Olympus Corporation, Monolith, Tokyo, Japan).
Molecular identification of antagonistic fungi and phylogenetic construction
-
To identify the isolate potential antagonistic fungi, the DNA was extracted by CTAB method[18,19]. The specific primers of ITS1/4 was used to amplify the sequences. The ITS sequence was amplified according to the above PCR procedure, and the products were detected by electrophoresis and sent to the biological engineer for sequencing. All sequences from this study were submitted to GenBank. BLASTn algorithm was used to compare with the GenBank sequence database. According to the results of BLASTn analysis, the ITS sequences of the top ten species with the highest similarity were included in the phylogenetic analysis as references. Use MEGA5.0's default settings for its multiple sequences and manually optimize to achieve maximum sequence similarity. A phylogenetic tree was constructed based on the neighbor-joining method. Bootstrap analysis was carried out using the p-distance model. A Bootstrap value of 1,000 repetitions was used to assess clade stability and phylogenetic tree reliability.
Antifungal activity analysis
Dual culture tests
-
All fungal strains were transferred from stored cultures onto PDA plates and cultured at 25 °C in the dark. A mycelial plug (5 mm in diameter) of Colletotrichum camelliae cut from the margin of the medium area was placed on the center of a new PDA plate (9 cm in diameter). In the tested group, the potential antagonistic fungi were inoculated at a distance of 4 cm from the pathogenic fungi with two 5 mm mycelial plugs on its left and right side, while the corresponding site was replaced by two 5 mm PDA plugs in the control group. Each combination had at least three biological replicates. All plates were cultured at 25 °C in the dark and observed daily. When the pathogenic fungi stopped growing or the colony spread over the whole plant in the control group, the antagonistic type and the area of the pathogen colony were recorded and measured. If the tested fungi inhibited the growth of the pathogen and grew faster than the pathogen or the parasitized pathogen, the fungi was considered to have an antagonistic effect against the pathogen in vitro.
Disease resistance experiments on tea leaves
-
The fungi with antagonistic effects in vitro were cultured in PDA medium in the dark at 25 °C. The mycelium disk with a diameter of 5 mm was penetrated from the edge of the actively growing colony. The disc was inoculated to potato glucose broth (PDB) and shake cultured (140 rpm) at 25 °C for 3 to 4 d, and spores were collected by centrifugation[21]. Then, the spore suspension was prepared to 104/ml, 105/ml, and 106/ml, respectively. Spore suspensions of different concentrations were sprayed on both sides of healthy tea leaves and incubated at 25 °C and 70% relative humidity for 48 h in the artificial climate chamber. The blank control (CK) was treated similarly, but sterile water was used instead of spore suspension. Each leaf was punctured with a sterile needle to create a symmetrical wound. Then, a 5 mm mycelial plug of pathogen was inoculated on the wound site at the back of the leaf. All leaves were cultured at 25 °C and 70% relative humidity for 48 h in the artificial climate chamber. The four time points of 1, 3, 5, and 7 d were observed, and the area of lesions was measured using ImageJ software.
The determination of antioxidant enzymes' activity
-
The UV-1800 spectrophotometer (Shimadzu, Japan) was used to determine the activity of superoxide dismutase (SOD), peroxidase (POD) and catalase (CAT) according to a previously described protocol[22]. In brief, the samples for the assessment of antioxidant enzymes, SOD, POD, and CAT were treated with nitrogen blue tetrazole, guaiacol, and hydrogen peroxide methods, respectively.
Statistical analysis
-
Statistical analyses were performed using Excel and GraphPad Prism 9. The means and standard deviations of the data were calculated and statistically analyzed with analysis of t-test.
-
To identify epiphytic fungi that exhibit antagonistic effects against tea plant anthracnose pathogens, epiphytic fungi from healthy tea plant leaves were initially isolated and purified. These fungi were then tested for pathogenicity and assessed for their antagonistic properties. The findings revealed that a total of 13 epiphytic fungi were purified, which belonged to various genera, including Cercospora, Cladosporium tenuissimum, Fusarium oxysporum, Nemania bipapillata, Phoma sp., Phytophthora, Trichoderma, Aspergillus niger, Macrocybe gigantea, and Talaromyces (Table 1). Among them, nine species were not pathogenic to tea leaves. We conducted plate antagonism experiments on these nine epiphytic fungi against the pathogen Colletotrichum camelliae, respectively. As a result, it was found that four of them exhibited antagonistic properties against pathogens (Fig. 1). Furthermore, En10 and En11 exhibit antagonistic behavior towards pathogens through types of substrate competition and mycoparasitism, whereas En12 and En13 exhibit antibiosis type (Table 1, Fig. 1). Therefore, the En10 and En12 strains were selected for further analysis due to their unique antagonistic characteristics. Through examination of colony and spore morphology (Fig. 2a & b) and phylogenetic identification (Fig. 2c), it was determined that En10 and En12 were affiliated with Aspergillus niger and Talaromyces, respectively. These results suggest that these two endophytic fungi may have potential as biocontrol agents against tea plant anthracnose pathogen.
Table 1. Information of endophytes isolated from tea plant.
No. Cultivar Taxonomy Pathogenicity Antagonistic type En01 'Shaancha1' Cercospora No − En02 'Shaancha1' Cercospora Yes − En03 'Shaancha1' Cercospora Yes − En04 'Shaancha1' Cladosporium tenuissimum No − En05 'Shaancha1' Fusarium oxysporum Yes − En06 'Shaancha1' Nemania bipapillata Yes − En07 'Shaancha1' Phoma sp. No − En08 'Shaancha1' Phytophthora No − En09 'Shaancha1' Trichoderma No − En10 'Longjing 43' Aspergillus niger No AC En11 'Longjing 43' Macrocybe gigantea No AC En12 'Longjing 43' Talaromyces No B En13 'Longjing 43' Talaromyces No B In column of 'Antagonistic Type', A, B and C represent competition for substrate, antibiosis and mycoparasitism, respectively. 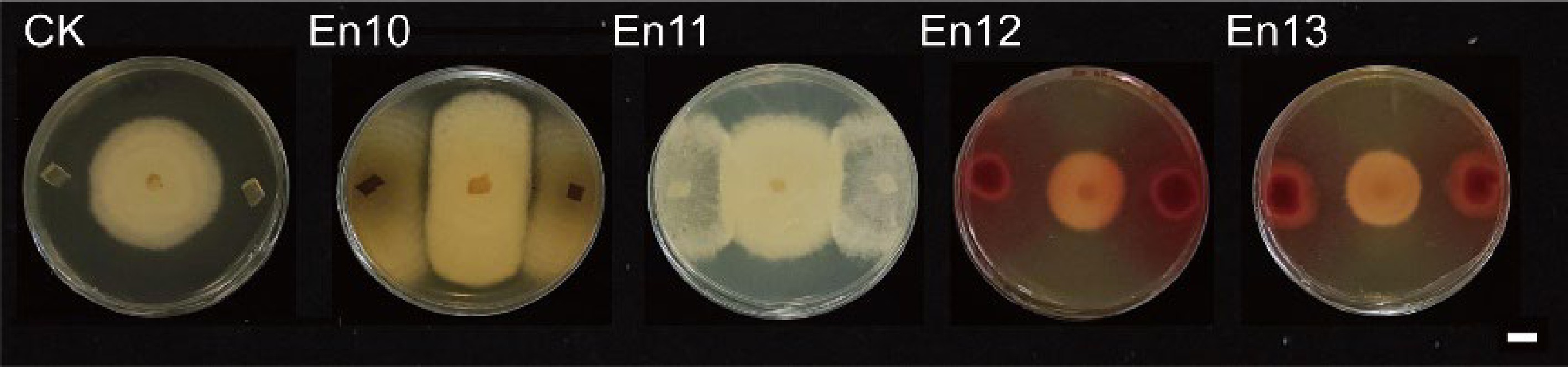
Figure 1.
Plate antagonism experiments on endophytic fungi against the pathogen Colletotrichum camelliae. Scale bar = 1 cm. CK: Two 5 mm PDA plugs were used as controls.
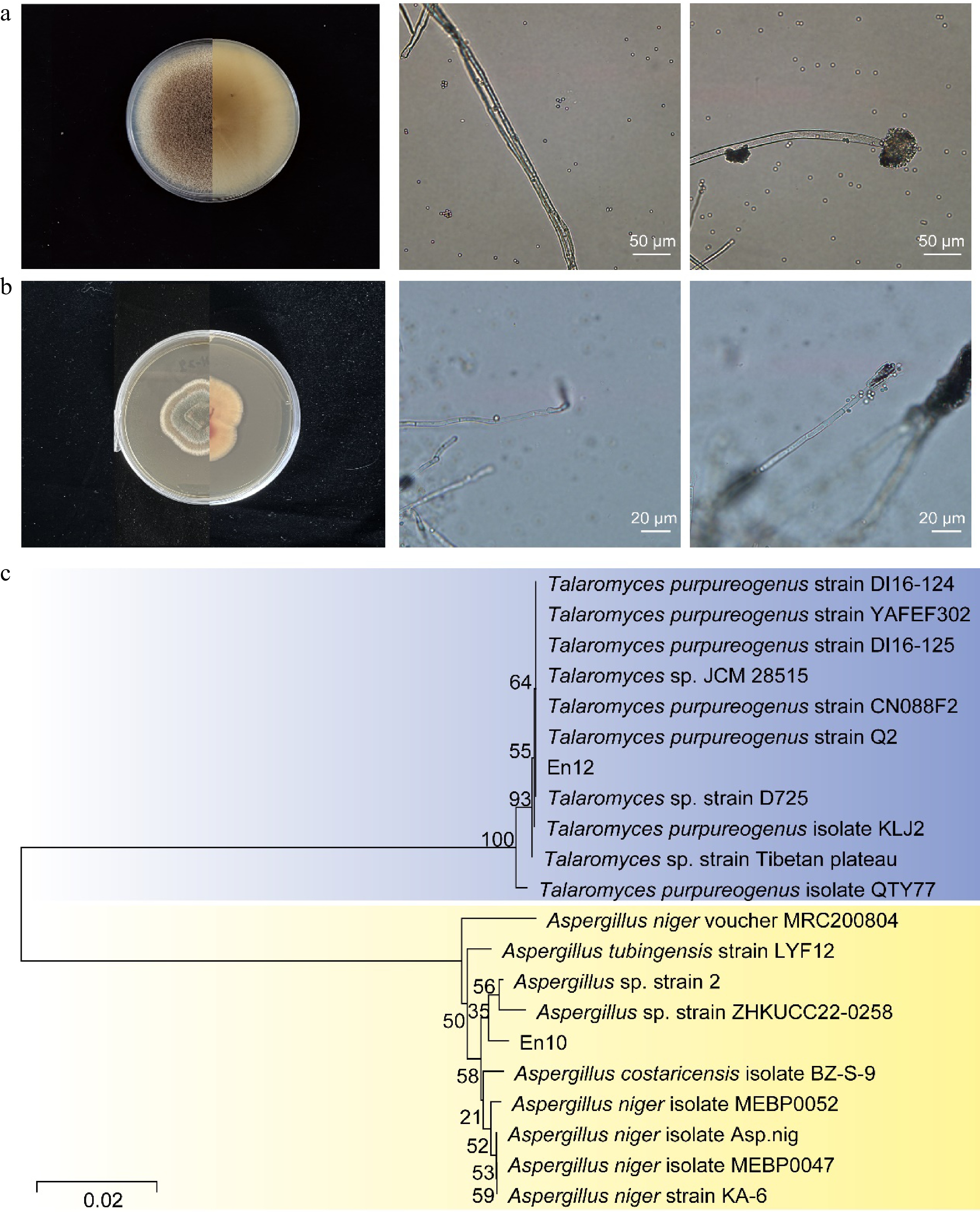
Figure 2.
Morphological and molecular identification of En10 and En12. (a) Front and reverse view of En10 colony on PDA after 9 d, displaying hyphae and conidia. (b) Front and reverse view of En12 colony on PDA after 9 d, showing hyphae and conidia. (c) Phylogenetic analysis based on sequences of the internal transcribed spacer between En10 and En12.
Inhibition of colony growth of anthracnose pathogen by antagonistic fungi
-
To further verify the inhibition of antagonistic fungi against the anthracnose pathogen, a dual culture test was executed. The results demonstrated that the two antagonistic fungi had a significant inhibitory effect on the colony growth of the anthracnose pathogen. After a culture period of 9 d, En10 and En12 exhibited inhibitory rates of 59% and 44% on the growth of the anthracnose pathogen, respectively (Fig 3). These results further confirm the antagonistic effects of En10 and En12 against the tea plant anthracnose pathogen in vitro.
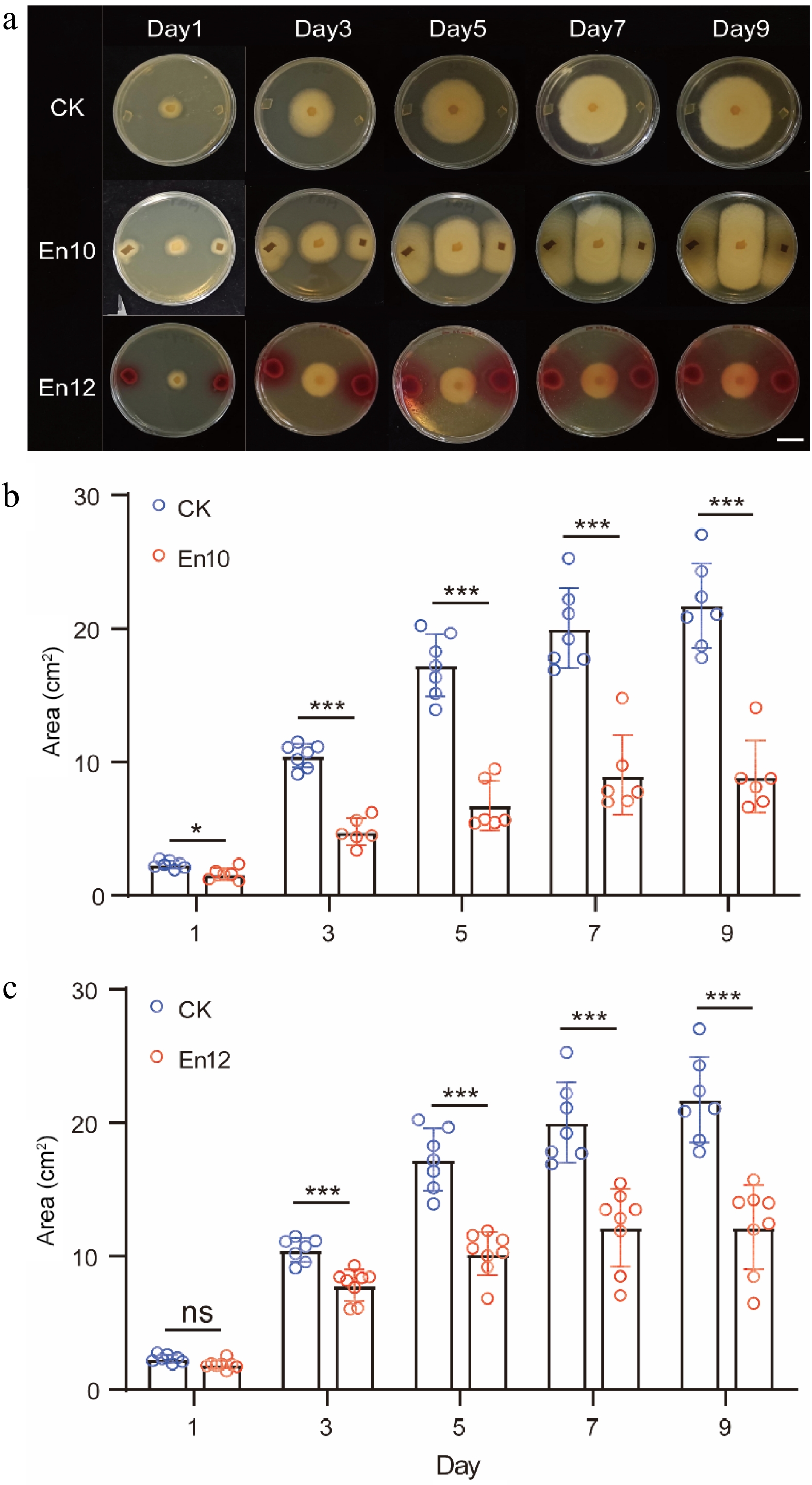
Figure 3.
Inhibition of colony growth of anthracnose pathogen by antagonistic fungi En10 and En12. (a) Dual culture test on anta gonistic fungi against the anthracnose pathogen. CK: Two 5 mm PDA plugs were used as controls. Scale bar = 1 cm. (b), (c) Area change of pathogen under the condition of antagonistic culture. The data represent the average ± SD of biological repeats, ns represents no significant difference and * stands for significant difference analysis.
Control effect of antagonistic fungi against anthracnose on tea leaf
-
To further elucidate the efficacy of antagonistic fungi in controlling anthracnose in tea leaves, we conducted resistance experiments on tea plant. Tea leaves were sprayed with spore suspensions of antagonistic fungi at three distinct concentrations, including 104/ml, 105/ml and 106/ml. However, it was observed that spraying high and medium concentrations of spore suspension followed by inoculation pathogen resulted in a significant number of tea leaf deaths. For instance, after spraying 106/ml concentration of En10 and En12, the mortality rate of tea leaves reached 14.29% and 66.70% on the third day, respectively, while the control group had a mortality rate of 0%. On the fifth day, the mortality rate of tea leaves reached 42.86% and 83.30%, respectively, while the control group had a mortality rate of 42.86% (data not shown). Therefore, the concentration of 104/ml spore suspension was chosen for treatment. The results of the study demonstrated a noteworthy reduction in the incidence rate of tea leaves that were treated with antagonistic fungi, as opposed to the control group. This suggests that the application of antagonistic fungi on tea leaf has the potential to inhibit tea plant anthracnose (Fig. 4a & b).
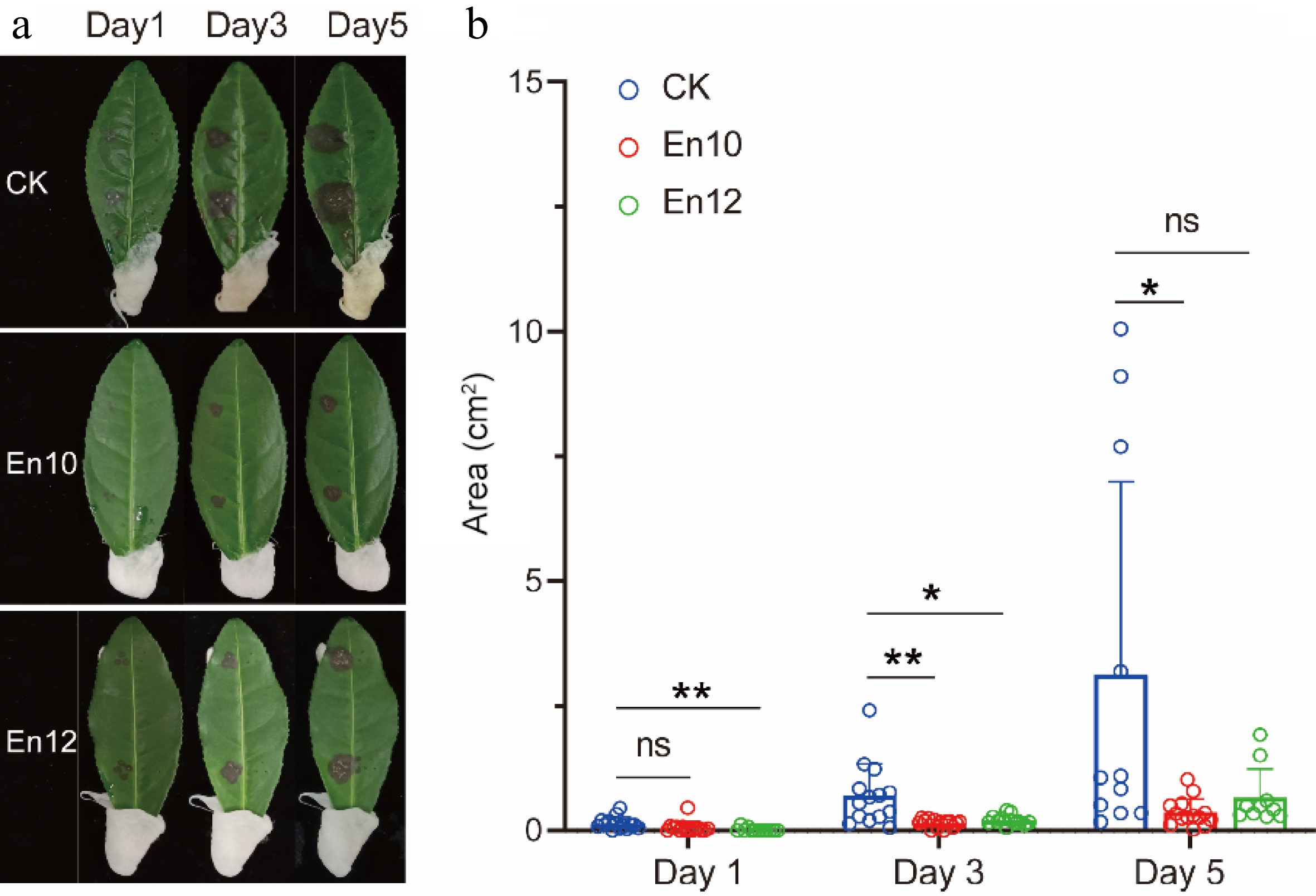
Figure 4.
Determination of the control effect of antagonistic fungi against the anthracnose on tea leaves. (a) and (b) indicate the images and the diseased area of tea leaves treated by blank solvent, fungi spores of En10 and En12 after 1, 3, and 5 d, respectively. The data represent the average ± SD of biological repeats. * means significant difference (p ≤ 0.05); ** represents significant difference (p ≤ 0.01); ns indicates no significant difference.
Activities of antioxidant enzymes after En10 and En12 treatment
-
The activities of antioxidant enzymes in tea leaves treated with En10 and En12 were measured. The results indicated a significant upregulation of SOD and POD enzyme activities following En12 treatment compared to the control (Fig. 5a & b). Particularly noteworthy was the 5.47-fold increase in SOD activity, while CAT enzyme activity remained unchanged (Fig. 5c). In contrast, although En10 exhibited notable resistance to the anthracnose pathogen, treatment of tea leaves with it did not result in significant alterations in the activity of associated antioxidant enzymes (Fig. 5). It is suggested that En12 may enhance the antioxidant response of tea leaves, increasing their resistance to anthracnose.
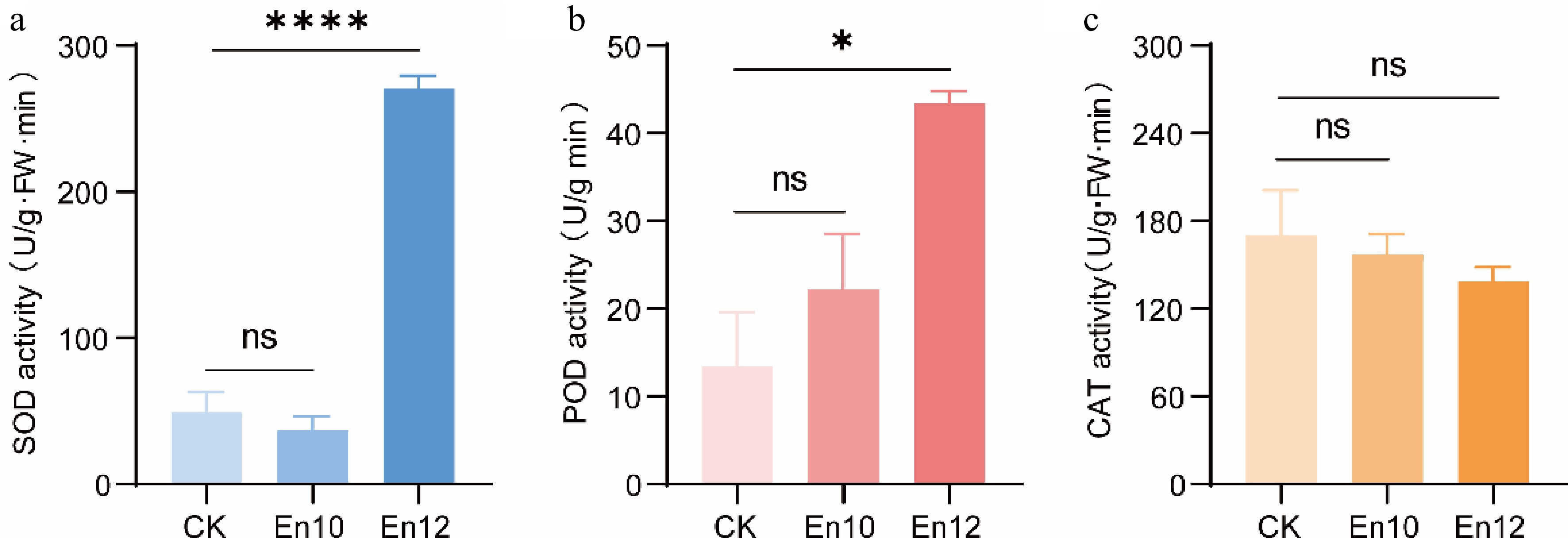
Figure 5.
Detection of enzyme activity in tea leaves after spraying with antagonistic fungi. (a)−(c) indicates the SOD activity, POD activity and CAT activity, respectively. CK: sterile water treated. En10 (En12): antagonistic fungi spores treated. The data represent the average ± SD of three biological repeats. * stands for significant difference analysis and ns indicates no significant difference.
-
Anthracnose, a prevalent disease affecting tea plant worldwide, hinders plant growth and diminishes tea quality[2]. Current control methods rely heavily on chemical agents. However, increasing consumer demand for healthier products has highlighted concerns over pesticide residues in tea and environmental contamination in tea gardens. The emerging field of endophytes in plant defense against pathogens presents promising alternatives for combating fungal, bacterial, viral, and pest-related threats in tea cultivation[17]. The potential application of endophytic antagonism in managing tea plant diseases is promising due to the abundance of epiphytic fungi in tea plant. Therefore, researchers have isolated and identified endophytes from the tea plant. So far, endophytes known to exhibit antagonistic activity against anthracnose include Bacillus amylolyticus[7], Bacillus subtilis[23], Bacillus atrophaeus[24], Bacillus velezensis[25] and Streptomyces luteosporeus[26]. In this study, epiphytic fungi were isolated from the leaves of healthy tea leaves, resulting in the purification of 13 epiphytic fungi representing Cercospora, Cladosporium tenuissimum, Fusarium oxysporum, Nemania bipapillata, Phoma sp., Phytophthora, Trichoderma, Aspergillus niger, Macrocybe gigantea, and Talaromyces (Table 1). Notably, nine of these species were found to be non-pathogenic to tea leaves. Experimental findings demonstrated significant antagonistic effects against anthracnose pathogens by two specific epiphytic fungi, En10 and En12 (Fig. 1). Subsequent identification revealed that En10 and En12 were classified as Aspergillus niger and Talaromyces, respectively (Fig. 2). This represents the first documented instance of their antagonistic activity against tea plant anthracnose.
Aspergillus niger is a common species of Aspergillus fungi, which is widely found in food, plant products, and soil species[27]. It has a strong ability to produce enzymes, and can produce a variety of high-activity extracellular enzymes, such as cellulase, pectinase, amylase, acid protease, glucose oxidase, and so on. Therefore, it is widely used in the fermentation industry. Some studies have shown that adding Aspergillus niger during tea fermentation can promote the decomposition of tea protein and affect the quality of tea[28,29]. In this study, it was found that Aspergillus niger En10 had a significant inhibi tory effect on the growth of anthracnose pathogen colonies (Figs 1 & 3). After spraying 104/ml spore suspension of En10 on healthy tea leaves (Fig. 4), the incidence of anthracnose inoculation was significantly reduced. This suggests that Aspergillus niger En10 has potential as a biocontrol agent against tea anthracnose.
Talaromyces are important decomposers in nature and some Talaromyces fungi can produce important enzymes. For example, Talaromyces verruculosus, Talaromyces pinophilus and Talaromyces purpureogenus can produce large amounts of cellulase and xylanase[30−33]. Some metabolites of Talaromyces can also enhance the absorption of minerals by plants, thereby improving the disease resistance and stress resistance of plants. A strain of Talaromyces En12 was identified, which could significantly improve the resistance of tea leaves to anthracnose after spraying a spore suspension (Fig. 4). Furthermore, the antagonistic activity of En12 may be attributed to its enhancement of the antioxidant capability of tea leaves, such as the activities of SOD and POD (Fig. 5). This finding highlights the potential of using epiphytic fungi, such as Aspergillus niger and Talaromyces, to be developed as low-cost biocontrol agents for effectively controlling tea plant anthracnose.
Studies have shown that endophytes can improve plant resistance to pathogens through different mechanisms, such as producing secondary metabolites to inhibit the growth of pathogens and inducing plant defense mechanisms. Endophytes can also induce the production of polyphenol oxidase, peroxidase, phenylalanine ammoniase, and superoxide dismutase in plants to improve the disease resistance of host plants. Prabhukarthikeyan et al. found that after treatment by Pseudomonas fluorescens, the enzyme activities of POD, PPO, PAL, SOD and CAT in roots and stalks of turmeric have been significantly improved, and they have effectively prevented and controlled the root rot disease of turmeric[34]. Khan et al. isolated from the roots of tomato a penicillium strain LK5[35]. Its secondary metabolites can increase the activity of POD, CAT, PPO, and GSH, to improve the defense capability of the plant itself. In our investigation, significant alterations in SOD and POD enzyme activities in tea leaves were observed following treatment with En12 (Fig. 5). These changes potentially contribute to enhancing the tea plant's resistance against anthracnose pathogen incursion. Nonetheless, our primary focus centered on the screening and characterization of epiphytic fungi with efficacy against tea anthracnose. Further research is warranted to elucidate the mechanisms of epiphytic fungi action, isolate and identify antibacterial constituents, and explore the interactions between epiphytic fungi and their host tea plant. Overall, our findings demonstrate the potential of using epiphytic fungi as a cost-effective and environmentally friendly approach for controlling tea plant anthracnose.
-
Thirteen strains of epiphytic fungi were isolated and purified from healthy tea leaves, with nine found to be non-pathogenic. Through plate confrontation tests, two strains, En10 identified as Aspergillus niger, and En12 identified as Talaromyces, were observed to significantly inhibit the growth of pathogens. Dual culture experiments and disease resistance trials on tea leaves demonstrated that En10 and En12 effectively suppressed the invasion of anthracnose disease. Additionally, En12 displayed potential to improve the antioxidant response of tea leaves, thereby increasing resistance to anthracnose. In all, this study identified two epiphytic fungi, belonging to Aspergillus niger and Talaromyces, that have the potential to be used as biological control agents for the management of tea plant anthracnose.
-
The authors confirm contribution to the paper as follows: study conception and design: Wang W, Yu Y, Bao L; tea samples collection: Liu L, Jiao M, Ma Z, Ma Y, Zhou J; experiment conduction: Guan H, Liu L, Xie X, Bao Y; data analysis, visualization: Guan H, Liu L, Wang W; original draft preparation: Wang W, Liu L. All authors reviewed the results and approved the final version of the manuscript.
-
All data supporting the conclusions of this study may be found in the publication, which are available online. Any additional relevant information can be obtained from the corresponding author upon request.
This work was supported by the National Natural Science Foundation of China (32202551), China Agriculture Research System of MOF and MARA (CARS-19) and the National key research and development program of China (2022YFD1602003). Some texts in this paper were polished by Stork's Writing Assistant (
www.storkapp.me/writeassistant ).-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Lu Liu, Haonan Guan
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu L, Guan H, Jiao M, Ma Z, Bao Y, et al. 2024. Identification of two antagonistic fungi and antifungal activity analysis against anthracnose in tea plant (Camellia sinensis). Beverage Plant Research 4: e032 doi: 10.48130/bpr-0024-0020 

Identification of two antagonistic fungi and antifungal activity analysis against anthracnose in tea plant (Camellia sinensis)
- Received: 22 March 2024
- Revised: 28 April 2024
- Accepted: 13 May 2024
- Published online: 03 September 2024
Abstract: Anthracnose, a prevalent disease affecting tea leaves, poses a significant threat to tea yield and quality. Current control measures predominantly rely on chemical pesticides, raising concerns over environmental contamination and pesticide residues. In exploring biological alternatives, 13 epiphytic fungi were isolated from healthy tea leaves, with nine demonstrating non-pathogenic characteristics. Through a plate confrontation test, strains exhibiting high antagonistic activity against anthracnose were identified, notably En10 and En12, which effectively suppressed the growth of tea plant anthracnose pathogens. Morphological and molecular analyses classified En10 as Aspergillus niger and En12 as Talaromyces. The application of spore suspensions of varying concentrations of En10 and En12 onto tea leaves revealed a significant reduction in anthracnose incidence. Notably, En12 exhibited the capacity to enhance the antioxidant potential of tea leaves. These findings underscore the potential of utilizing antagonistic fungi En10 and En12 for anthracnose control in tea plant, offering valuable insights and technical foundations for biological management strategies against this disease.
-
Key words:
- Tea anthracnose /
- Epiphytic fungi /
- Aspergillus niger /
- Talaromyces /
- Biological control