









-
Perilla frutescens (L.) Britton, belonging to Lamiaceae, is a variety of versatile economic crop, also commonly used in herbal medicine. It has been widely cultivated in China, Japan, South Korea and many other Asian countries in recent years[1]. The leaves of Perilla are used for the preparation of cold granules in Traditional Chinese Medicine (TCM), and as vegetables and spices in Asian countries[1,2]. The leaves of Perilla possess various chemicals, including terpenes, flavonoids, phenolic acids, etc.[3]. The medicinal value of Perilla has been officialized in the Chinese Pharmacopoeia and the catalog of affinal drugs and diet[1,4]. The essential oils of Perilla leaves are the major medicinal flavor components. They are also widely applied in the skin care and aromatization industry[5,6].
The essential oils of Perilla leaves include an abundant diversity of chemical types, which are classified into monoterpene (MT)-type oils and phenylpropene (PP)-type oils[7,8]. Interestingly, there are multiple kinds of monoterpenes in the leaves of Perilla genus, which can be further classified into the following six chemotypes according to their principal constituents: perillaldehyde (PA), perillaketone (PK), perillene (PL), piperitenone (PT), citral (C) and elsholtziaketone (EK) types[9]. Among these monoterpenes, PA are the major aromatic medical ingredient for prescriptions in China and Japan, while PK were thought to be a potent lung toxin[10]. Besides these main monoterpenes, geraniol (GL) is an acyclic monoterpene also commonly found in a wide range of Perilla plants[11], and linalool (LL) can be found in all Perilla plants and maybe a dead-end compound in general monoterpene biosynthetic pathways[12]. Wherein, PA, PL, GL, and LL are commercially important for perfumery, food and medicine[13,14]. As multiple chemical types of monoterpenes are enriched in Perilla, Perilla hence has been considered to be a model system for the study of monoterpene metabolism.
The biosynthesis of monoterpene is specially localized to the glandular trichomes and initiated from the mevalonate (MAV)/methylerythritol 4-phosphate (MEP) pathway in plants[15−17]. Then the terpene synthases (TPS) catalyze prenyl pyrophosphates, the products of MVA and MEP pathways, to the formation of terpene compounds, and cytochrome P450s (CYP450s) further modify the backbones of these terpenes[17]. Recently, some TPSs and CYP450s involved in Perilla monoterpene biosynthesis have been reported, including limonene synthase, GL synthase, LL synthase, and mono-oxygenase[18−20]. Enzymes participated in PA biosynthesis, such as CYP71D18 and CYP17A7146, have been characterized[18−20] . Eight double-bond reductases (PfDBRs) that catalyze the conversion of isopyrone and soyone to PK were identified by enzymatic methods in vitro[21]. The recent high-quality and chromosome-scale Perilla genome data establishes a solid foundation for the characterization of Perilla monoterpene biosynthesis[22]. In the present study, transcriptome analysis was carried out on four different Perilla chemotypes some TPSs involved in monoterpene biosynthesis were identified, as well as multiple regulatory factors responsible for this biosynthesis pathway. Furthermore, the function of selected TPSs were characterized by a heterologous expression system and in vitro enzyme assay. These results collectively will help to understand the molecular mechanism of Perilla monoterpene biosynthesis and analyze the biosynthetic pathways of terpenes in Perilla.
-
The Perilla cultivars of four chemotypes, including PA-, PK-, PL- and PT-types, were selected and planted in the greenhouse of Guangzhou University of Chinese Medicine (Guangzhou, Guangdong, China, 113°41' E; 23°07' N). The PA-type cultivar, with purple wrinkled leaves, belongs to P. frutescens var. crispa. The PK-, PL- and PT-type cultivars, with green non-wrinkled leaves, belong to P. frutescens var. frutescens (Fig. 1a). The leaves at seedling stages were collected for gas chromatography-mass spectrometry (GC-MS) analysis and RNA extraction. All samples are stored in liquid nitrogen immediately after sampling.
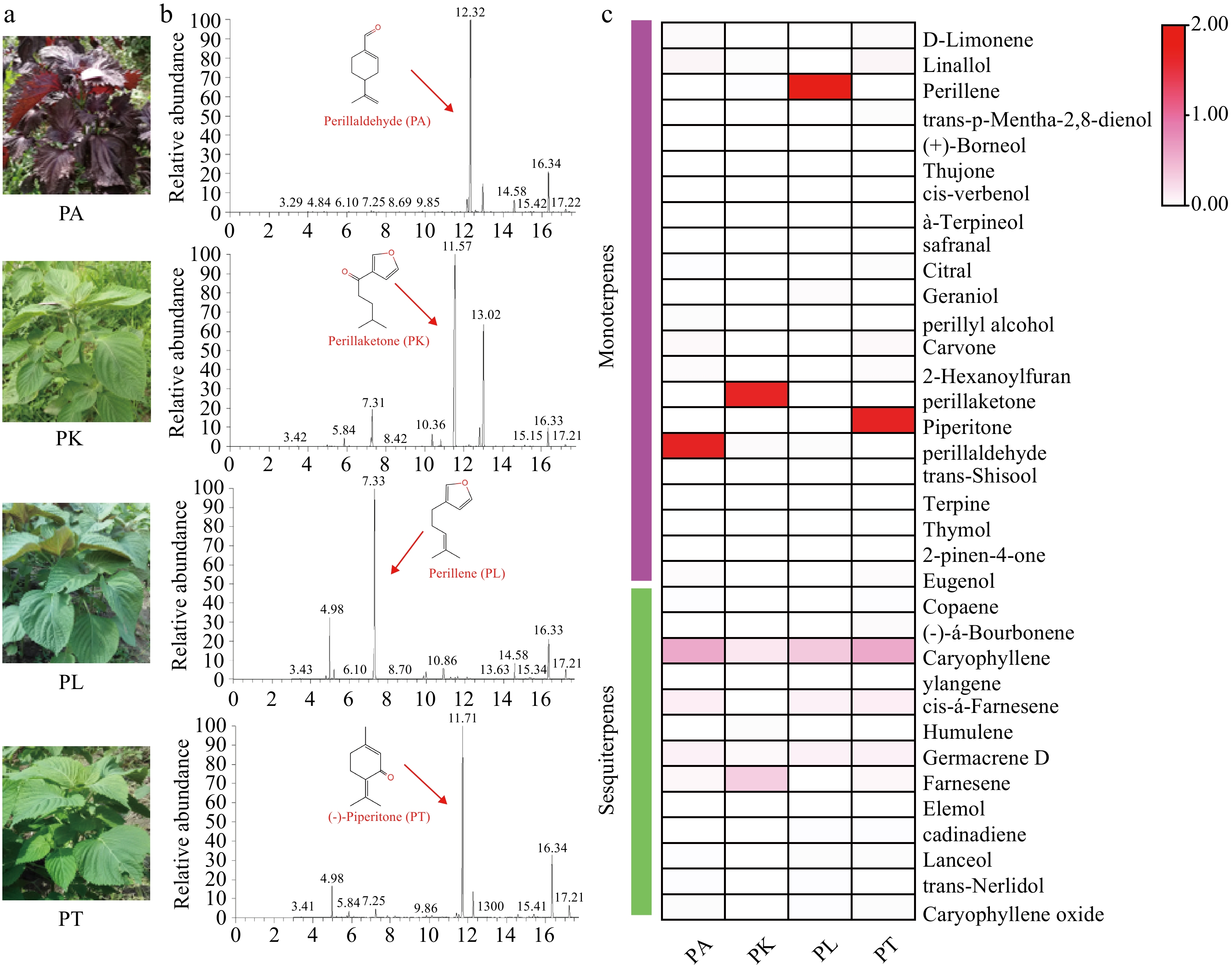
Figure 1.
The analysis of volatile components in four Perilla cultivars. (a) Phenotype of four chemical types of Perilla; (b) GC-MS analysis of volatile essential components from four Perilla leaves; (c) Heatmap of metabolite contents in four Perilla leaves.
GC-MS analysis
-
Perilla leaves of four chemotypes were crushed. Then, 0.2 g leaves powder was extracted by petroleum ether and filtered for GC-MS analysis. Analysis conditions include: RXT-5 MS quartz capillary column (30 m × 0.25 μm × 0.25 μm); The front column pressure is 63.9 kPa; The initial temperature of the column was 80 °C and was retained for 1 min. After the heating rate of 15 °C/min, the column was increased to 300 °C and retained for 15 min. MS conditions: ionization mode EI, filament current 0.5 mA; Electron energy 70 eV; Multiplier voltage 0.86 kV; Ion source 230 °C, solvent delay 1 min; Plasma/nucleus ratio m/z: 40~500. The NIST spectrum library was retrieved by Agilent qualitative software, and the chemical structure analysis was combined to identify the species of components. The relative percentage of each chemical component of volatile oil was calculated by the peak area normalization method.
Transcriptome analysis
-
Total RNA from 12 Perilla leaves were extracted using the RNApre Pure plant RNA extraction kit (DP432) (Tiangen, Beijing, China). The mRNA sequencing library was constructed using the NEBNext® Ultra RNA Library Prep Kit (New England Biolabs Inc., Ipswich, MA, USA). Then, the sequencing library was analyzed using the Agilent 2100 Bioanalyzer and was sequenced by an Illumina HiSeq™ 2000 sequencing system (Illumina Inc., San Diego, CA, USA). The original transcriptome data has been uploaded to the NGDC database (National Genomics Data Center) (Number: PRJCA021059).
The Perilla genome data came from the National Center for Biotechnology Information (NCBI, Accession No.: GCA_019511825.2)[22]. Trimmomatic software is used for quality control of transcriptome data[23]. STAR(v2.7.10a) software was used to build an index and clean data was compared to the Perilla genome[24]. The Python module HTseq is used for P. frutescens transcriptome data quantification[25]. Gene expression levels of fragments per kilobase of transcript per million mapped reads (FPKM) were calculated. Then differentially expressed genes (DEGs) were identified using DESeq2[26]. Genes with |log2 (Fold change) |≥ 1| and p < 0.01 were considered DEGs. The online tool eggnog (
http://eggnog-mapper.embl.de ) was used to annotate the whole genome protein of Perilla[27]. R package ClusterProfiler was used for GO (Gene ontology) and KEGG (Kyoto Encyclopedia of Genes and Genomes) enrichment analysis of differential genes[28].Identification and characteristic analysis of metabolosynthetic genes
-
The two hidden Markov models (HMM) of TPS (Terpene_synthase, PF01397 and Terpene_synthase_C, PF03936) were downloaded from Pfam (
http://pfam.xfam.org/ ) and the Perilla genome was searched[29]. The identified PfTPS proteins were further determined by online HMMER (www.ebi.ac.uk/Tools/hmmer ) and a phylogenetic tree was constructed using the Neighbor-Joining method in MEGA X (Bootstrap 1000)[30]. Heatmaps of candidate genes were drawn using TBtools (v1.112) (https://github.com/CJ-Chen/TBtools )[31]. Intergenomic collinearity analysis using MCScanX[32]. Chromosomal localization and collinearity results were visualized using TBtools. According to the Annotation information of Metabolic pathway synthase in KEGG and eggnog, the encoded gens were identified in the MVA/MEP pathways in Perilla. The Python script is used to calculate the correlation coefficient between genes expression, using the Pearson correlation coefficient[33].Real-time PCR analysis and cloning of related genes
-
The full-length transcripts of PfTPSs genes were derived by 5' RACE-PCR and/or 3' RACE-PCR. Then the PCR products were ligated to the PLB vector (Tiangen) and sequenced by Sangon Biotech. Primer3Plus (
www.primer3plus.com/index.html ) was used to design primers for PfTPS genes. The primers used for genes cloning were listed in Supplemental Table S1. The fluorescence quantitative reaction system was referred to Wu et al., three replicates were used in each group, and PfActin was used as the key gene for analysis[34]. For data analysis, refer to the 2−ΔΔCᴛ method[35].Functional characterization in heterologous expression system
-
All the cloned PfTPSs genes were introduced into the heterologous expression vector pETDuet-1. Then the expression plasmids were transformed into C41 Escherichia coli (E. coli) strain. The positive colony were firstly cultivated in TB medium at 37 °C to an initial OD600 of 0.4-0.6, and then the cultures were induced by 1 mM IPTG for another incubation of 72 h at 16 °C. After the cultivation, the cultures were extracted for three times by equal volume of n-hexane, and then the extracts were concentrated by rotary evaporation instrument for gas chromatography-mass spectrometry (GC-MS) detection.
In vitro enzymatic reaction
-
The selected PfTPSs genes were introduced into pET28a vector and transformed into E. coli strain BL21 (DE3). The positive colony were incubated in LB medium at 37 °C to the initial OD600 of 0.4−0.6, and then the cultures were induced by 0.5 mM IPTG for another 12 h of cultivation. The cultured cells were harvested and resuspended in lysis buffer (50 mM Tris-HCl, 500 mM NaCl, 20 mM imidazole, 20 mM β-mercaptoethanol, pH 8.0) for 30 min at 4 °C. Then the cells were disrupted by ultrasonication and the lysate was centrifuged at 13,000 g and 4 °C for 30 min. The crude proteins were inside the supernatant. For the soluble PfTPSs proteins, the crude proteins were purified by HIS nickel column.
For in vitro enzymatic reaction, crude or purified PfTPSs proteins were added to the reaction buffer containing 200 mM Tris-HCl (pH 7.5), 40 mM MgCl2, 10% glycerol and 1 mM geranyl pyrophosphate (GPP) as the precursor. After incubation at 30 °C for 30 min, the reaction system was extracted using an equal volume of n-hexane and then detected by GC-MS.
-
The volatile essential oils components of leaves from four Perilla cultivars were analyzed using GC-MS analysis (Fig. 1a, b). A total of 35 terpenes, including 22 monoterpenes and 13 sesquiterpenes were detected in these cultivars. The main monoterpenes are consistent with their chemotypes classification. 68.01% of PA was identified as the main compounds in PA-type cultivars, while 71.65% of PL, 88.76% of PK and, 61.20% of PT are the main compounds in PL-type, PK-type, PT-type varieties, respectively (Fig. 1b). The GL (0.07%−0.68%) and LL (0.03%−1.77%) were the ubiquitous and trace metabolites that existed in these Perilla cultivars. Besides these main chemicals, other monoterpenes and sesquiterpenes were also identified, including limonene, borneol, thujone, verbenol, citral, carvone, trans-shisool, terpine, thymol 2-pinen-4-on, caryophyllene, germacrene, farnesene, trans-nerlidol etc. (Fig. 1c; Supplemental Table S2).
Transcriptome analysis
-
To explore the molecular mechanism involved in different monoterpenes biosynthesis, the transcriptome analysis was performed for the leaves of four chemotype cultivars. After sequence and data filtration, a total of 579 million clean reads, comprising 86.90 Gb nucleotide bases with an average 46.26% GC were obtained (Supplemental Table S3). The average 98% clean reads were assembled to the Perilla genome (GCA_019511825.2) (Supplemental Table S4). Then, the gene annotation and differential expression analysis were carried out among the four cultivars. The PA type are the main medicinal component of Perilla according to Chinese Pharmacopoeia. Hence, more attention is focused on the PA-type. In total, 236 specific up-regulated genes were compared with other three cultivars (Fig. 2a). In the specific up-regulated genes of PA-type, phylpropanoid, and monoterpene biosynthesis were enriched using the KEGG enrichment analysis (Fig. 2b; Supplemental Table S5). Meanwhile, the comparison analysis among the other three different chemotype cultivars, the 79, 92 and 155 specific up-regulated genes were identified in PK-type, PL-type, and PT-type, respectively (Fig. 2c, e & g). Genes involved in terpenes, including monoterpene, sesquiterpene, diterpene and triterpene were enriched in the corresponding chemical type (Fig. 2d−f; Supplemental Tables S6−S8).
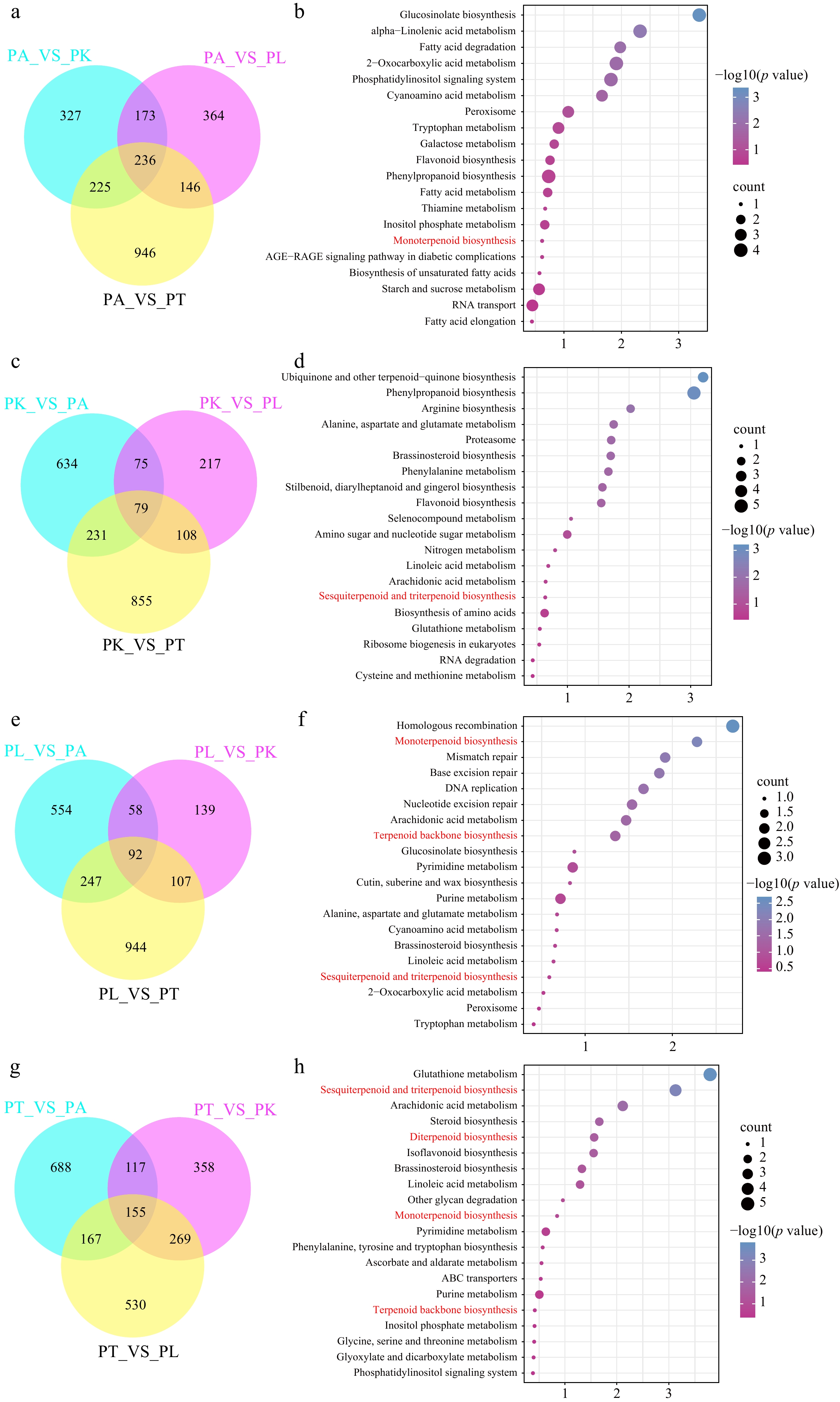
Figure 2.
Different expression genes and KEGG enrichment analysis of four Perilla cultivars. (a), (c), (e), (g) The intersection of PA, PL, PK, PT4 chemotypes with the other three chemotypes are indicated in the Venn diagrams. (b), (d), (f), (h) KEGG enrichment analysis of special up-expressed genes in PA, PL, PK, PT-type, respectively.
Biosynthetic pathway of the volatile oils in P. frutescens
-
The MEP and MVA pathways are the basic terpene biosynthesis pathways. The 69 genes encoding 17 enzymes in MEP and MVA pathways were identified in the four cultivars (Fig. 3; Supplemental Table S9). The MEP pathway starts with pyruvate, which is catalyzed by DXS to form 1-deoxy-D-xylulose-5-phosphate. Subsequently, it is continuously catalyzed by DXR, MCT, CMK, MDS, HDS, and HDR to form MEcPP. There were two encoded genes of DXR, MCT, CMK, and MDS in Perilla which showed upregulated expression in PA-type. In the MVA pathway, Acetyl-CoA is catalyzed by AACT HMGS, HMGR, MVK, PMK, MPDC IPP, and DMAPP to IPP. Finally, the equilibrium between IPP and DMAPP are controlled by isopentenyl diphosphate isomerase (IPPI)-encoded genes and the further reaction synthesize by geranyl pyrophosphate synthase (GPPS)-encoded genes to produce the GPP in plastids (Fig. 3; Supplemental Table S10).
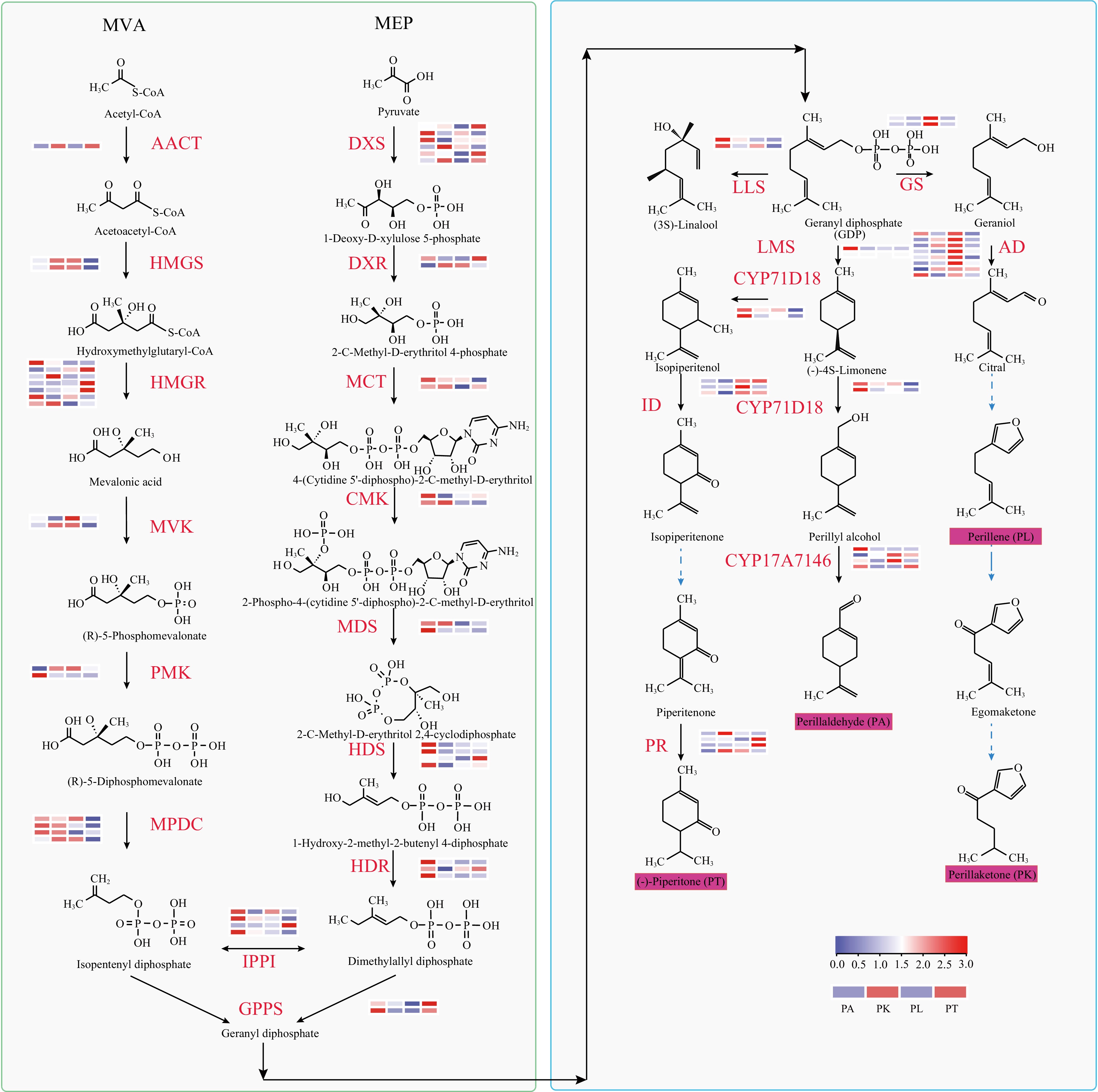
Figure 3.
Synthesis pathway and single thread synthesis pathway of P. frutescens isoprene. The MEP pathway: 1-deoxy-D-xylulose 5-phosphate synthase (DXS); 1-deoxy-D-xylulose-5-phosphate reductoisomerase (DXR); 2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase (MCT); 4 diphosphocytidyl-2-C-methyl-D-erythritol kinase (CMK); 2-C-methyl-D-erythritol 2,4-cyclodiphosphate synthase (MDS/MECPS); (E)-4-hydroxy-3 methylbut-2-enyl-diphosphate synthase (HDS); 4-hydroxy-3-methylbut-2-en-1-yl diphosphate reductase (HDR); The MVA pathway: acetyl-CoA C-acetyltransferase (AACT); hydroxymethylglutaryl-CoA synthase (HMGS); hydroxymethylglutaryl-CoA reductase (HMGR); mevalonate kinase (MVK); phosphomevalonate kinase (PMK); diphosphomevalonate decarboxylase (MPDC); isopentenyl-diphosphate Delta-isomerase (IPPI); geranyl diphosphate synthase (GPPS); Linalool synthase (LLS); Geraniol synthase(GS); limonene synthase (LMS); alcohol dehydrogenase (AD); Cytochrome P450 proteins (CYP); isopiperitenol dehydrogenase (ID); pulegone reductase (PR).
Moreover, the biosynthesis pathway of PA and PT has been reported in Perilla and Mentha, respectively[18−20,36]. Limonene is the common substrate for the synthesis of PA and PT. Two genes encoded limonene synthase (LMS) were identified in Perilla. For the biosynthesis of PA, limonene is catalyzed by CYP71D18 and CYP17A7146. Two genes encoded CYP71D18 were found and up-expressed in PA-type, while four paralogs encoded CYP17A7146 were identified and one of them showed upregulated expression in PA-type (Fig. 3; Supplemental Table S11). The biosynthesis of PT by isopiperitenol dehydrogenase (ID) and pulegone reductase (PR) were identified in Perilla, which possesses three and four encoded genes and shows different expression in four cultivars, respectively. (Fig. 3; Supplemental Table S11. Moreover, geraniol is catalyzed by GL synthase (GLS). Two genes encoded GLS and showed upregulated expression in PL. GL were further catalyzed by alcohol dehydrogenase (AD) to produce citral, which is the precursor of PL and PK. Besides them, LL synthase (LLS) are also the common monoterpene compounds in Perilla. Two encoded genes encoded LLS and up-expressed in PA-type were identified in Perilla (Fig. 3).
Identification and characterization analysis of the TPS family
-
The TPS use prenyl pyrophosphates as the substrate to synthesize terpenes, which are important for various chemotype formation in Perilla. In Perilla, a total of 109 TPS family members were identified using HMM search. The putative PfTPS proteins ranged from 230 to 817 amino acids in length (Supplemental Table S12), with the exon number from 3 to 15 (Fig. 4b). All members contained N-terminal (PF01397) and C-terminal (PF03936) conserved domains of TPS genes (Fig. 4c), and the RRX8W domain existed in the N-terminal, while the typical DDXXD conserved domain, as well as the typical functional domain RXR, existed in the C-terminal (Fig. 4d). The PfTPS genes displayed different expression trends in the four chemotypes (Fig. 4e).
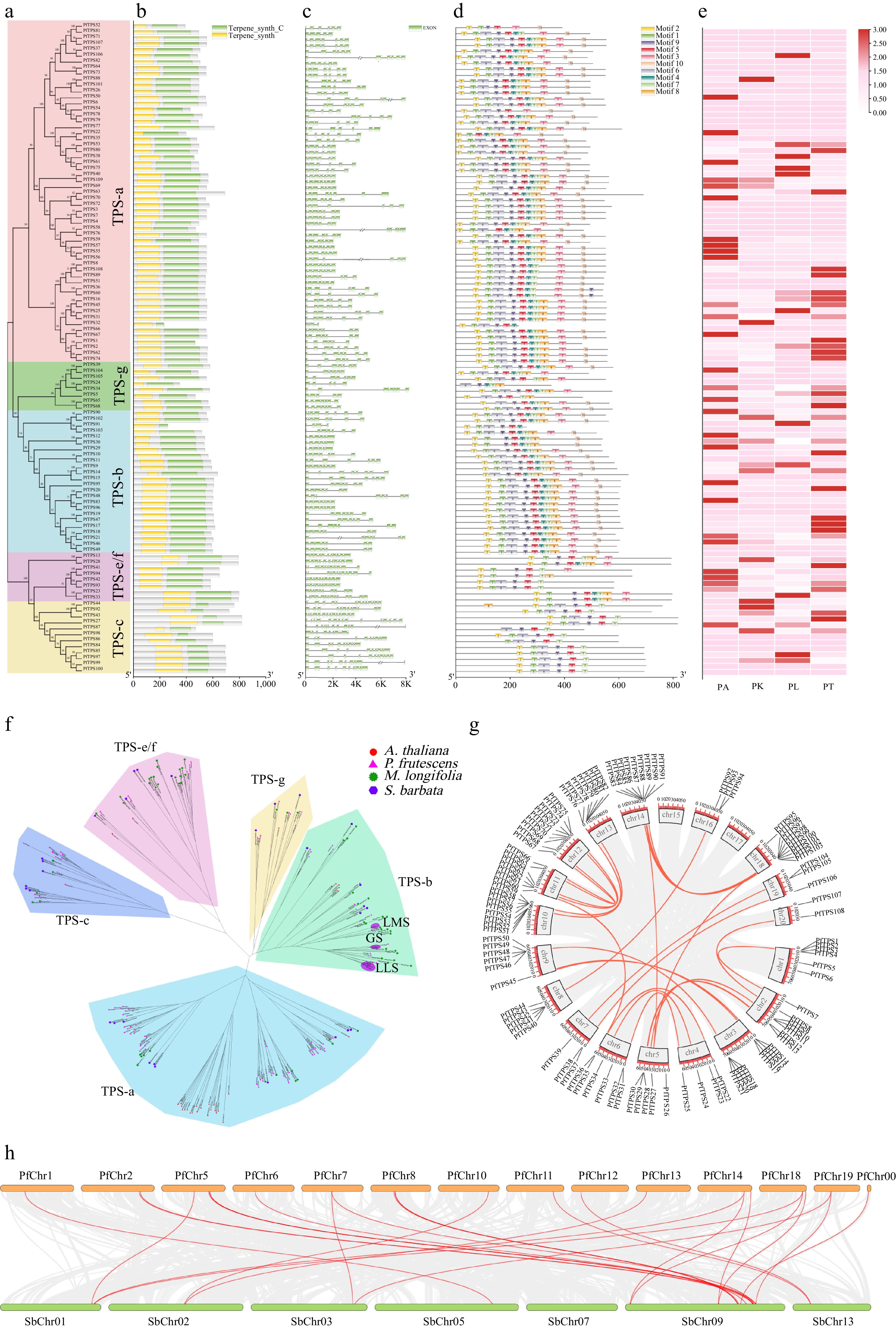
Figure 4.
The PfTPS Gene Family Characteristics in P. frutescens. (a)−(e) Phylogenetic evolutionary tree, conservative domain, gene structure, protein motifs and expression heatmap of PfTPS genes. (f) Subfamily classification of the PfTPS Family (LLS: linalool synthase; GS: geraniol synthase; LMS: limonene synthase). (g) Chromosomal localization and collinearity analysis of Perilla. (h) Collinearity analysis of TPS in Perilla and S. baicalensis.
PfTPS family members were divided into five major subfamilies, including TPS-a (57 members), TPS-b (24 members), TPS-c (12 members), TPS-e/f (eight members), and TPS-g (eight members) (Fig. 4f). The number of TPS genes in Perilla (109) showed significant expansion compared to that of Arabidopsis thaliana[37], Solanum lycopersicum[38], and other lamiaceae species, including Mentha longifolia[39], Salvia miltiorrhiza[39], Ocimum tenuiflorum[39], and Lavandula angustifolialabiaceae[40]. Among the TPS in Perilla, the TPS-a and TPS-b accounts for the 57.29% and 22.02% proportion as main expanded sub-families in Perilla. (Supplemental Table S13). To explore the evolutionary relationship of TPS, chromosome mapping, and collinear block analysis were carried out. The PfTPS genes were unevenly distributed on the 18 chromosomes. As the tetraploid genome of Perilla, the distribution of allele genes in pairs is a normal phenomenon. There were nine PfTPS genes found on chromosome Chr10/11/12/13, 5/8 PfTPS in Chr5/12, and 4/6 PfTPS in Chr4/6, which showed obvious collinearity in the Perilla genome (Fig. 4g). The collinearity relationship between Perilla and S. baicalensis was further analyzed. Further analysis of the collinear relation between Perilla and S. baicalensis was performed. The collinear block in 11 Perilla chromosomes correspond with seven chromosomes in S. baicalensis. Universally, the TPS in Perilla showed tandem duplication, containing 45 indicating that there is an obvious evolutionary relationship between TPS of Perilla and S. baicalensis. However, it is especially that SbChr09 has obvious chromosome blocks corresponding with multiple chromosomes of Perilla (Fig. 4h).
Co-expression analysis and verification of functional TPS genes
-
To further mine the functional TPS genes in various chemotypes, the gene co-expression analysis was performed. Interestingly, PfTPS18, PfTPS46, PfTPS47, and PfTPS49 as significant core genes were identified. To present the significant relationship between those TPSs, the co-expression network was present in the core TPS and the terpene biosynthesis genes and TFs, respectively. Firstly, PfTPS18 as core genes were significant co-expression with 201 the terpene biosynthesis genes and TFs, including GPPS, HMGS, HDR, AACT, and ERF, MYB, NAC etc. (Fig. 5a). Similarly, PfTPS46 and PfTPS47 were also co-expressed with IPPI, HDR, GPPS, FPPS, HMGS, and MPDC genes, which are important rate-limiting genes in the MVA/MEP biosynthesis pathway (Fig. 5b, c). PfTPS49 was co-expressed with other five PfTPSs, including PfTPS15, PfTPS24, PfTPS38, PfTPS39, and PfTPS63, and associated with The GPPS, HMGS, and other TFs (Fig. 5d).

Figure 5.
The co-expression analysis and verification of PfTPSs. (a)−(d) Co-expression analysis with core PfTPSs, including (a) PfTPS18, (b) PfTPS46, (c) PfTPS47 , (d) PfTPS49 and five other PfTPSs. (e) qRT-PCR verification of PfTPS genes and MVA pathway genes (qRT-PCR results (left, line) and transcription results (right, bar)).
Among them, 12 PfTPS genes and two MVA pathway genes were selected for expression verification using qRT-PCR. The significant coincident gene levels were identified in transcriptome sequencing and qRT-PCR (r > 0.9). Those PfTPSs present general transcription in four cultivars but showed high expression in one chemotype. Such as PfTPS18, PfTPS21, and PfTPS76 showed up-regulated expression in PA types, especially PfTPS49 presents a specific high expression level. PfTPS46, PfTPS47, and PfTPS62 showed up-regulated expression in PL types. The expression levels of PfTPS87, PfTPS93, and PfTPS108 were similar in the four chemical types (Fig. 5e).
Functional characterization of PfTPSs from different chemotypes of Perilla
-
As PfTPS18, PfTPS46, PfTPS47, and PfTPS49 were the significant core genes according to the co-expression analysis, we selected these PfTPSs genes for further functional characterization. Due to the expression levels of the above PfTPSs in different chemotypes (Fig. 5e), genes with predominant expression levels in specific chemotypes were the only successful clones, such as the highest-expressed PfTPS18 and specific-expressed PfTPS49 in PA-type. Thus, the cloned PfTPSs genes were named with their chemotypes as follows: PfTPS46-PL, PfTPS46-PK, PfTPS18-PA, PfTPS47-PA, and PfTPS49-PA, respectively. To predict the possible catalytic functions of these PfTPSs, phylogenic analysis was performed.
To identify the catalytic functions of the cloned PfTPSs, the CDSs of different TPS to the expression vectors were ligated and transformed into E. coli to characterize the functions of PfTPSs. After being cultivated for 3 d, the cultures were extracted by n-hexane and then the compounds were detected by GC-MS analysis. Strain with PfTPS46-PL produced one peak in GC-MS profile compared to the control group (strain with control vector) (Fig. 6a). The product was determined to be linalool by the comparison of the retention time in total ion chromatograms and the mass spectrum with authentic standard linalool (Fig. 6a; Supplemental Fig. S1a, S1b). As the signal peptide (SP) region in the N terminal of PfTPS46-PL might affect the catalytic activity of the enzyme inside E. coli cells, this region was removed in the CDS of PfTPS46-PL and its function explored using the truncated variant. The strains harboring the truncated PfTPS46-PL brought the same linalool product in GC-MS analysis (Fig. 6a; Supplemental Fig. S1c). Next, to further confirm the catalytic function of PfTPS46-PL, the purification of the PfTPS46-PL protein was attempted and its function characterized using an in vitro enzymatic reaction. The proteins of PfTPS46-PL were not obtained due to its insolubility. Thus, the crude proteins of PfTPS46-PL were used with geranyl pyrophosphate (GPP) as precursor. Consistent with the result in the heterologous expression system, the crude PfTPS46-PL protein also produced the sole product linalool (Fig. 6b). According, PfTPS46-PL is a linalool synthase.
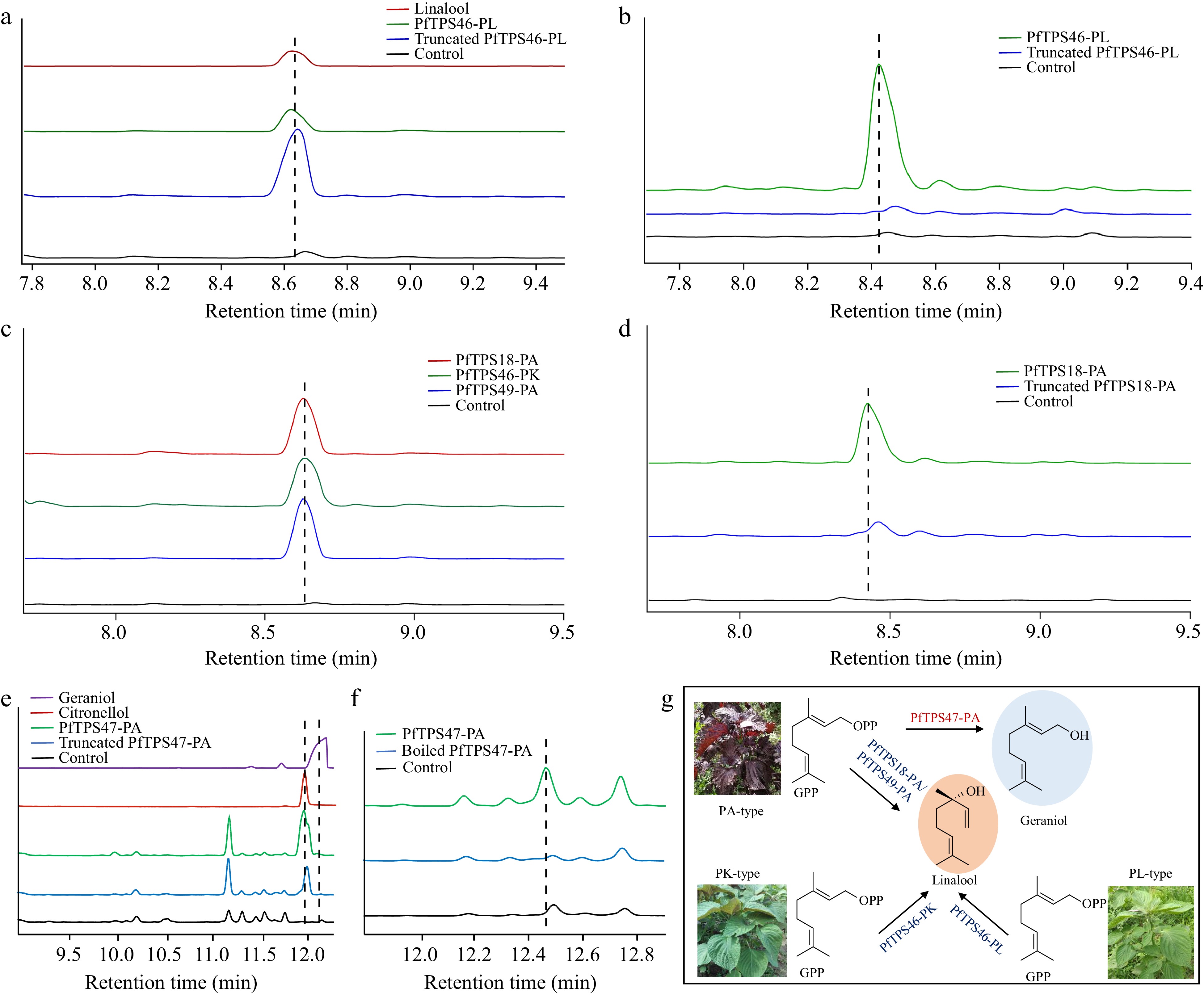
Figure 6.
Functional characterization of four PfTPSs. (a) Heterologous functional characterization of PfTPS46-PL and (b) the in vitro enzymatic reaction of PfTPS46-PL. (c) Heterologous functional characterization and (d) the in vitro enzymatic reaction of PfTPS46-PK, PfTPS18-PA, and PfTPS49-PA. (e) Heterologous functional characterization and (f) the in vitro enzymatic reaction of PfTPS47-PA. (g) Catalytic model of four PfTPSs.
Next, the functions of other candidate monoterpene synthases were characterized using the same strategy. The products of PfTPS46-PK, PfTPS18-PA, and PfTPS49-PA were all found to be linalool in the heterologous expression system, as well as the truncated enzymes (Fig. 6c; Supplemental Figs S2, S3). The crude proteins of PfTPS18-PA were selected as the representative for the in vitro enzymatic analysis. The crude proteins also produced the sole product linalool (Fig. 6d; Supplemental Fig. S4). The results indicated that these PfTPSs are linalool synthases.
For the function characterization of PfTPS47-PA, two products were detected in PfTPS47-PA and truncated PfTPS47-PA-harboring strains, with the major product citronellol and the minor product geraniol (Fig. 6e; Supplemental Figs S5, S6). However, the purified PfTPS47-PA protein catalyzed GPP to the sole product geraniol (Fig. 6f). Here, we speculated that some certain enzyme inside E. coli cells catalyzed geraniol, the product of PfTPS47-PA, to citronellol. The results showed that PfTPS47-PA was highly similar to geraniol synthase while other PfTPSs were assigned to the linalool synthase category. Collectively, we identified four linalool synthases and one geraniol synthase in different Perilla chemotypes (Fig. 6g).
-
The essential oils of Perilla are well-recognized aromatic compounds and possess multiple pharmacological effects. They are also the valuable genetic materials of monoterpene biosynthesis and regulation for multiple kinds of chemical types. In the present study, four monoterpene chemotype cultivars were selected. The important monoterpene biosynthesis pathway and important candidate TPSs were analyzed and verified using transcriptome sequence and heterologous expression verification.
Terpene biosynthesis initiates from the MVA and MEP pathway in plants[16]. Compared with Arabidopsis, the number of encoded genes in the MVA and MEP pathways increased significantly in Perilla. The gene amplification could induce gene differentiation and affect the biosynthesis of terpenes[38]. The genes encoded HMGS, PMK, and MDPC in the MVA pathway, DXR, MCT, CMK, and MDS in the MEP pathway, and IPPI, and FPPS in the cross-flow pathway were found to have obvious expansion in Perilla. Interestingly, most MEP pathway genes were up-regulated in the PA type, which implied the high-efficiency biosynthesis in the PA type of Perilla.
Various volatile oil components as chemical type of Perilla have been researched. In the early stage, the genetic basis for the monoterpene chemical type in Perilla was verified using artificial hybrids method. The chemical composition is controlled by a series of multiple alleles (G1, G2, g) and an independent pair of alleles (H, h)[41]. In the present research, 109 TPS members in Perilla genomes were identified. They also showed obvious gene expansion, especially, the expansion of TPS-a and TPS-b, reached 57.29% and 22.02% proportion, more than other majority of Lamiaceae plants. The expression and function of TPSs were also significantly differentiated. In the past few years, more geraniol synthases, linalool synthases, and limonene synthases have been acquired in Perilla[18−20]. The biosynthetic pathway of piperitenon in Mentha longifolia was also reported. Based on genome-wide identified, more GS, LLS, LMS, and the PA and PT biosynthesis encoded genes were also explored. Their expression trends are in accordance with volatile oil components. LMS and GS for example had high expression in PA-types and LLS had high expression in PL-types. Based on co-expression analysis, the four TPSs act as core genes in various chemical types. The high expression in PA types and PL types were selected for function verification.
The heterologous functional characterization and in vitro enzymatic reaction are two important methods for the functional characterization of TPSs. PfTPS18, PfTPS46, and PfTPS49 were characterized as linalool synthases and PfTPS47 was characterized as geraniol synthases, respectively. Linalool and geraniol are general compounds. The core genes in co-expression analysis were characterized as linalool and geraniol synthases. Further research will be carried out to identify more chemotype-related TPSs. Moreover, the genotype in different cultivars of a certain species is unique and widely used in the recognition of different cultivars in many plants (Supplemental Figs S7 & S8)[42,43]. For example, the polymorphic variant of one sesquiterpene synthase, VvTPS24, in grape conferred the cultivar a different product in the chemotype, which was distinguishable from other grape cultivars (Supplemental Fig. S9)[44]. However, the identified isozymes in different Perilla chemotypes, such as the linalool synthases, including PfTPS46-PL, and PfTPS46-PK, showed no obvious variations in their amino acid sequences. As the constitutions of compounds in different cultivars are determined by the enzymes and their expressions, we guess that the diverse content of linalool in different cultivars is caused by the regulatory elements.
-
The authors confirm contribution to the paper as follows: conceptualization, methodology: Yang SM, An TY, Shen Q; formal analysis: Yang SM, Wang YX, Lin GB; investigation: Yang SM, Wang YX, Lin GB, Chu HY; software: Yang SM, Chu HY, Wang YX; data curation, validation: Wang YX; supervision, writing – review & editing: An TY, Shen Q; resources, writing – original draft: Yang SM, Chu HY; funding acquisition: Shen Q. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets presented in this study are publicly available. RNA-seq data are available via NGDC with accession No. PRJCA021059.
This work was funded by the National Natural Science Foundation of China Grant (U22A20446) and the National Natural Science Foundation for Regional Fund (31860391).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Shi-Mei Yang, Hong-Ye Chu
- Supplemental Table S1 Real-time fluorescence quantitative primers.
- Supplemental Table S2 The volatile oil components of four perilla cultivars.
- Supplemental Table S3 Transcriptome sequencing data of four chemical leaves of Perilla.
- Supplemental Table S4 Mapping genome data of four chemical leaves of Perilla.
- Supplemental Table S5 The KEGG enrichment analysis involving in specific up-regulated in PA-type.
- Supplemental Table S6 The KEGG enrichment analysis involving in specific up-regulated in PK-type.
- Supplemental Table S7 The KEGG enrichment analysis involving in specific up-regulated in PL-type.
- Supplemental Table S8 The KEGG enrichment analysis involving in specific up-regulated in PT-type.
- Supplemental Table S9 Comparison of MVA and MEP pathway genes in Perilla and Arabidopsis.
- Supplemental Table S10 PA synthetic pathway genes.
- Supplemental Table S11 The gene and their expression level identification of biosynthesis pathway of perilla specific monoterpene.
- Supplemental Table S12 Characteristics of Perilla TPS gene family.
- Supplemental Table S13 TPS gene family characteristics in other species.
- Supplemental Fig. S1 The mass spectrum of linalool. A. The mass spectrum of standard linalool. B. The mass spectrum of the product of PfTPS46-PL. C. The mass spectrum of the product of truncated PfTPS46-PL.
- Supplemental Fig. S2 The mass spectrum of the product of PfTPS46-PK, PfTPS18-PA and PfTPS49-PA.
- Supplemental Fig. S3 The GC-MS analysis of the products of truncated PfTPS46-PK, PfTPS18-PA and PfTPS49-PA.
- Supplemental Fig. S4 The mass spectrum of the product of the crude proteins of PfTPS18-PA.
- Supplemental Fig. S5 The mass spectrum of citronellol. A. The mass spectrum of standard citronellol. B. The mass spectrum of the major product of PfTPS47-PA.
- Supplemental Fig. S6 The mass spectrum of geraniol. A. The mass spectrum of standard geraniol. B. The mass spectrum of the minor product of PfTPS47-PA.
- Supplemental Figs S7
- S8
- Supplemental Fig. S9
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yang SM, Chu HY, Wang YX, Guo BL, An TY, et al. 2024. Analysis of monoterpene biosynthesis and functional TPSs of Perilla frutescens based on transcriptome and metabolome. Medicinal Plant Biology 3: e017 doi: 10.48130/mpb-0024-0017 

Analysis of monoterpene biosynthesis and functional TPSs of Perilla frutescens based on transcriptome and metabolome
- Received: 02 February 2024
- Revised: 03 June 2024
- Accepted: 24 June 2024
- Published online: 03 September 2024
Abstract: Perilla is a characteristic medicinal and edible plant. According to the different monoterpenes in Perilla leaves, Perilla can be divided into many chemical types. In this study, four Perilla cultivars of different chemical types, including perillaldehyde (PA), perillaketone (PK), perillene (PL) and piperitenone (PT) were selected for metabolome and transcriptome analysis. In total, 35 terpene compounds were identified and main monoterpenes were consistent with corresponding chemical types, respectively. Combined with transcriptomes, 5,920 differentially expressed genes were obtained among them. In total, 69 genes referred to MVA (Mevalonate) and MEP (2-C-methyl-D-erythritol-4-phosphate) pathways and 109 PfTPSs encoded genes were identified, which possessed partial differentially expressed in four chemical types. The core PfTPSs in co-expression analysis were functionally characterized. Three PfTPSs were identified as linalool synthase and one PfTPSs was identified as geranyl synthase. This research analyzed the monoterpene biosynthesis and functional characterization of TPSs in Perilla, which can provide a foundation for in-depth research of the Perilla chemotype metabolic mechanism.
-
Key words:
- Perilla frutescens /
- Monoterpene /
- Chemotype /
- Terpene synthase /
- Functional characterization