









-
Tea plant (Camellia sinensis) is a widely cultivated perennial evergreen plant, which is used to produce health-beneficial tea beverages[1]. However, tea plant frequently suffers from pathogens due to the warm and humid growing environment, which leads to serious losses in yield and decreased quality in tea[2, 3]. Colletotrichum camelliae, as one of the dominant Colletotrichum species causing anthracnose in Ca. sinensis, caused large necrotic lesions in tea leaves and then resulted in the increased production losses[4]. In response to pathogen attacks, plants have evolved exquisite and effective defense systems[5]. The first tier of plant immune system is pathogen-associated molecular patterns (PAMPs)-triggered immunity (PTI), via the recognition of PAMPs by plant pathogen- or pattern-recognition receptors (PRRs)[6]. Pathogens can synthesize effectors and deliver them into the plant cell to counteract PTI, while plants can recognize the effectors via disease resistance proteins (R proteins) and implement effector-triggered immunity (ETI)[6, 7]. The majority of R proteins contain central nucleotide-binding site (NBS) and Apaf-1, R-protein, and CED-4 homology (ARC) subdomains, and leucine-rich repeats (LRRs) domain, called NBS-LRR or NLR receptors[8].
Plants use NBS-LRR proteins to perceive fungal pathogens by direct or indirect recognition of fungal effectors, triggering ETI and developing defense responses against pathogens[9, 10]. For example, the NBS-LRR proteins RGA4 and RGA5 in rice physically bind to two effectors ACR-CO39 and ACR-Pia of fungal pathogen Magnaporthe oryzae, resulting in resistance induction and the hypersensitive response (HR)[11, 12]. Over the years, the function of NLR genes in host plants has been reported. In tomato, one resistance (R) gene Sm encoding an NBS-LRR protein confers resistance to gray leaf spot disease caused by Stemphylium lycopersici[13]. In barley, virus-induced gene silencing of an NBS-LRR gene Rpg5 resulted in a compatible reaction with a normally incompatible stem rust pathogen Puccinia graminis, indicating the important role of Rpg5 as the stem rust resistance gene[14]. An NLR protein YrU1 elicits effective ETI after recognition of the effectors derived from the stripe rust fungus Puccinia striiformis f. sp. Tritici in bread wheat[10, 15]. Soybean Rps11 is an NLR gene conferring broad-spectrum resistance to Phytophthora sojae causing root and stem rot[16]. In apple, three NLR proteins MdRNL1, MdRNL2, and MdRNL3 contribute to the resistance to apple leaf spot caused by Alternaria alternate f. sp. mali[8]. Rice OsRLR1 gene encoding an NBS-LRR protein mediates resistance to the rice blast fungus M. oryzae through interaction with the transcription factor OsWRKY19[17]. Although the role of NLR genes in the resistance against fungal infection has been widely reported in many plants, few studies reported the contribution of NLR genes to disease defense in tea plant.
In our previous studies, 400 and 303 CsNLRs genes have been identified from the genomes of Ca. sinensis var. sinensis (CSS) and Ca. sinensis var. assamica (CSA), respectively. Based on the N-terminal domains, they were classified into two major groups, coiled-coil-containing NLRs (CNLs) and Toll/interleukin-1 receptor-containing NLRs (TNLs)[18]. The expression of these CsNLRs was induced by Colletotrichum, abiotic stresses, and exogenous methyl jasmonate (MeJA) by RNA-Seq analysis[18]. However, the role of CsNLRs in disease resistance have not been functionally validated. Here we identified one CsNLR gene, CsRPM1, whose expression was significantly induced by C. camelliae. Phylogenetic tree revealed that CsRPM1 was homologous to RPM1 in Arabidopsis, and clustered with CNL. It encodes a typical NBS-LRR protein located in the nucleus, cytoplasm, and cell membrane. Further functional analysis by transient expression confirmed that this gene confers resistance to C. camelliae in tea plant. The transcript level of CsRPM1 was induced by exogenous MeJA, and may be regulated by several transcription factors.
-
Five-year-old tea plant (Ca. sinensis cv. Longjing43) (LJ43) seedlings were grown under natural conditions. To analyze the expression of CsRPM1 at different stages of infection with C. camelliae, the tea leaves inoculated by C. camelliae strain LS_19 were sampled at 0, 3, 6, 12, 24, 48, 72, and 96 h post inoculation. To determine the expression of CsRPM1 under the induction of MeJA, tea leaves treated with exogenous 150 μM MeJA and then inoculated with C. camelliae strain LS_19 were sampled at 0, 24, 48, and 72 h after treatment. The collected samples were stored at -80°C for further experiments.
RNA-Seq analysis
-
The RNA-Seq expression data-set used to analyze the expression of CsNLRs in this study was obtained from our previous study, which was transcriptome of tea leaves in response to C. camelliae[19]. Differentially expressed genes (DEGs) were defined as their expression presented a > 1.5-fold change[18]. The heatmap was constructed by TBtools software[20].
qRT-PCR analysis
-
Total RNA was extracted from collected samples using FastPure® Plant Total RNA Isolation Kit (Polysaccharides& Polyphenolics-rich) (Vazyme Biotech Co., Ltd, China). cDNA was then synthesized using HiScript II 1st Strand cDNA Synthesis Kit (+gDNA wiper) (Vazyme Biotech Co., Ltd, China) according to the manufacturer's instructions. qRT-PCR assays were performed with ChamQ Universal SYBR qPCR Master Mix (Vazyme Biotech Co., Ltd, China) using the Bio-Rad CFX96TM Real-Time System (USA). CsPTB1 gene encoding polypyrimidine tract-binding protein was used as reference[21]. Primer pair (forward: 5'-TCTCCTTCGTCGCTTGTC-3' and reverse: 5'-ATAGGGTCTTCTGTTAGTCTGG-3') was used to amplify CsRPM1 for qRT-PCR. The experiment with at least three replicates was independently repeated three times.
Transient gene suppression of CsRPM1
-
To transiently inhibit the expression of CsRPM1 in LJ43, antisense oligodeoxynucleotide (asODN) was used. Candidate sequences of asODN were designed using Soligo (
http://sfold.wadsworth.org/cgi-bin/index.pl ) with CsRPM1 gene sequence as the input. The sense oligonucleotides (sODN) were used as the control. 10 sODNs-asODNs pairs were selected for synthesizing to ensure and improve the interference effect[22]. The synthesized 10 pairs of sODN and asODN were adjusted to the concentration of 30 µM with ddH2O and then injected into the tea leaves respectively. After incubation, the injected leaves were inoculated with the 5-mm mycelial plugs cut from C. camelliae strain LS_19 and then sampled at 12, 24, 48, and 72 h post inoculation. At each point in time, the lesions on tea leaves were observed and measured. The leaves samples were then harvested and stored at -80°C for further analyses. The experiment with at least three replicates was independently repeated three times.Subcellular localization of CsRPM1
-
The CsRPM1 coding sequence lacking the termination codon was amplified with the primers (forward: 5'-CGAGCTCGGTACCCGGGGATCCATGGCCTTGGCTGCCGTGGG-3' and reverse: 5'-CCTTGCTCACCATGGTGTCGACAGTCAATCCTGTGGAACGAG-3'), and then fused with the vector pCAMBIA2300-35S-eGFP using CloneExpress II One Step Cloning Kit (Vazyme Biotech Co., Ltd, China). The fused vector and control vector (pCAMBIA2300-35S-eGFP) were then introduced into Agrobacterium tumefaciens GV3101 using the liquid nitrogen quick-freezing method respectively. The activated A. tumefaciens containing the vectors (OD600 = 0.5) were infiltrated into 5-week-old Nicotiana benthamiana leaves (expressing nuclear marker-RFP)[23]. After two days in the dark, the GFP fluorescence signal was observed using laser confocal scanning microscopy (Zeiss LSM 780, Germany). Excitation wavelengths of RFP and GFP were 532 and 488 nm respectively.
Screening of candidate transcription factors (TFs) regulating CsRPM1 expression
-
TeaCoN website (
http://teacon.wchoda.com/ ) was used for candidate TFs screening with CsRPM1 as the query[24].Statistical analysis
-
Data were the mean ± standard deviation of three biological replicates. The mean and standard errors were respectively calculated using the MEAN and STDEV function in Excel 2019. Statistically significant differences were determined with one-way ANOVA analysis (*, p < 0.05; **, p < 0.01) via SPSS Statistics 18 software.
-
To identify and analyze the expression pattern of DEGs in LJ43 inoculated with C. camelliae, we performed RNA-Seq analysis. Results from Venn diagram identified 12 DEGs at different stages of infection (12, 24, and 72 h) (Fig. 1a). Notably, among the 12 DEGs, there was an NLR-encoding gene (TEA004876) that was significantly up-regulated at 72 h compared to the control group (Fig. 1b). TEA004876 was predicted as the disease resistance protein RPM1, and could be induced under various biotic and abiotic stresses (
http://tpdbtmp.shengxin.ren:81/analyses_search_locus.html?id=TEA004876 ). It was named CsRPM1, and selected for further characterization. Phylogenetic analysis showed that CsRPM1 was clustered with RPM1 belonging to CNL (CC-NBS-LRR) group (Fig. 1c). CsRPM1 gene in LJ43 encodes an NBS-LRR protein, containing a signal peptide, a NB-ARC domain, and multiple LRR motifs (Fig. 1d).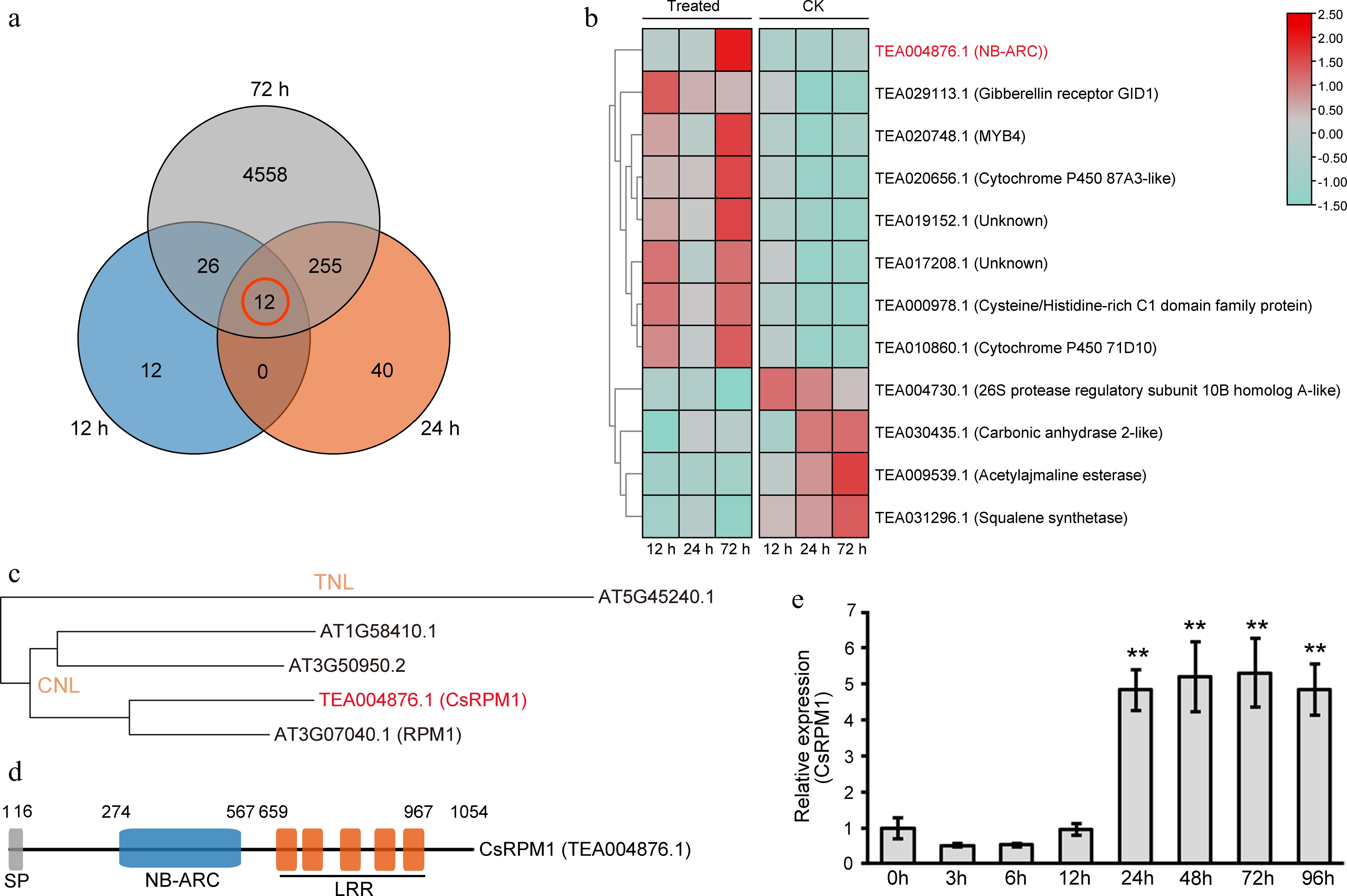
Figure 1.
Identification and expression pattern of CsRPM1 in tea plant leaves inoculated with C. camelliae. (a) Venn diagram of DEGs in tea plant leaves inoculated with C. camelliae at 12, 24, and 72 h. (b) Heat maps of the expression of 12 DEGs in LJ43 at 12, 24, and 72 h after inoculation with ddH2O (Control) or C. camelliae (Treated). (c) Phylogenetic tree constructed via the TNL and CNL genes. (d) Domain map of CsRPM1 protein. The domain prediction of CsRPM1 was performed with SMART analysis service. (e) The expression pattern of CsRPM1 in tea plant leaves during different stages of infection with C. camelliae by qRT-PCR analysis. **, p < 0.01.
The expression pattern of CsRPM1 at different stages of infection with C. camelliae was further confirmed by qRT-PCR analysis. The result showed that the expression of CsRPM1 was significantly up-regulated at 24, 48, 72 h, and 96 h after inoculation (Fig. 1e), suggesting that CsRPM1 can be induced by the infection with C. camelliae and probably involved in disease resistance in tea plant.
Silencing of CsRPM1 impairs host resistance to C. camelliae
-
To further valid the role of CsRPM1 in disease resistance in tea plant, asODN was used to transiently inhibit the expression of CsRPM1 in LJ43. The transcript of CsRPM1 was significantly down-regulated at 12 and 72 h post injection with 30 µM asODN compare to the control group (transformed with sODN) (Fig. 2a). At 72 h, the necrotic lesions caused by C. camelliae on the asODN group were significantly larger than those on the sODN control group (Fig. 2b, c). The results indicated that silencing of CsRPM1 facilitates the infection by C. camelliae and impairs the resistance of tea plant to C. camelliae.
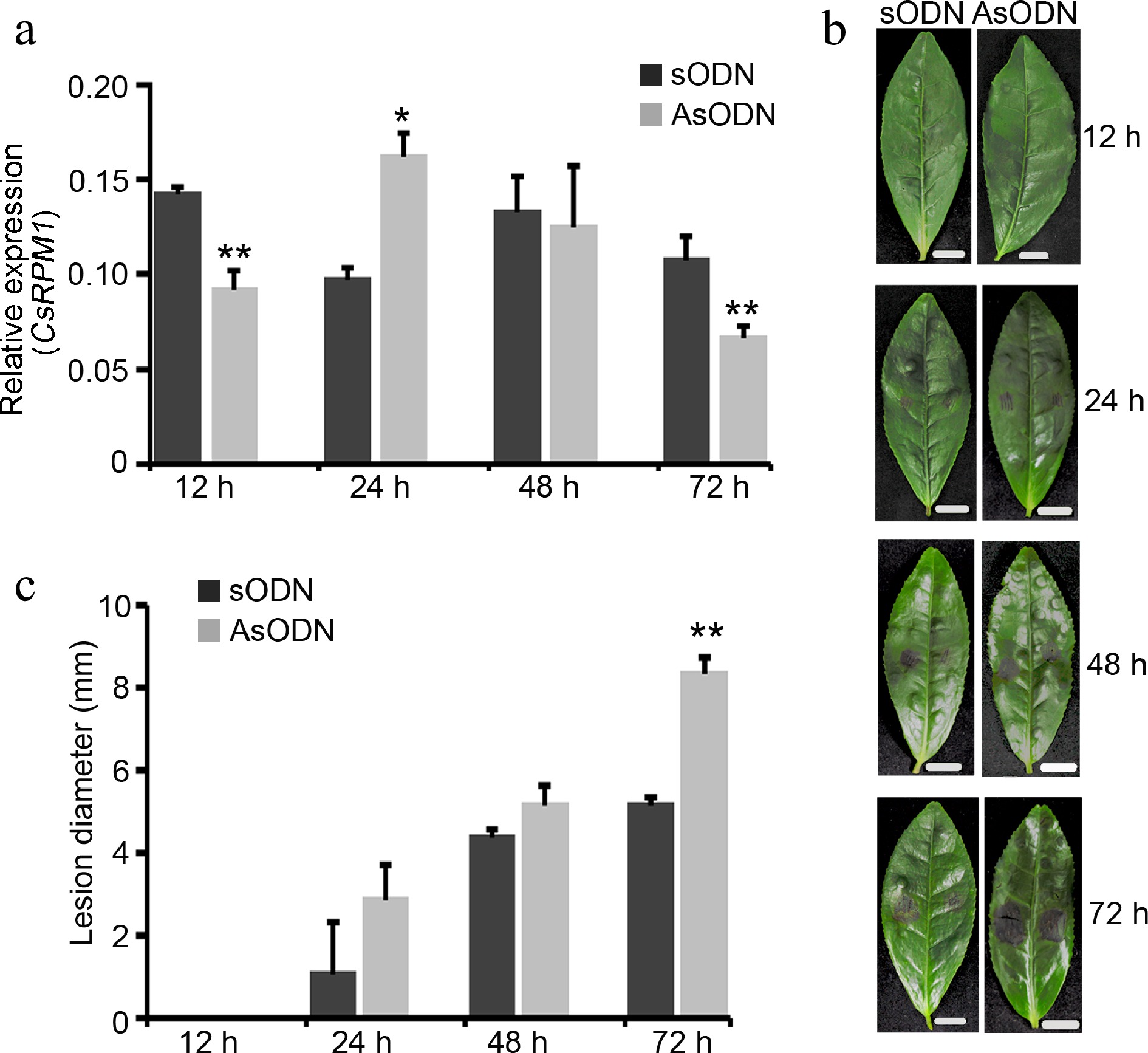
Figure 2.
Silencing of CsRPM1 impairs host resistance to C. camelliae. (a) Expression level of CsRPM1 gene at different stages after injection with 30 µM asODN or sODN. *, p < 0.05; **, p < 0.01. (b) Symptoms in detached tea leaves that were injected with 30 µM asODN or sODN caused by C. camelliae strain LS_19. Bar = 0.5 cm. (c) Bar chart showing statistical analysis of the sizes of necrotic lesions in (b). Error bars represent standard deviation. **, p < 0.01.
Subcellular localization of CsRPM1
-
For examining the localization of CsRPM1 protein, we constructed the fused vectors expressing CsRPM1-GFP under the 35S promoter. When CsRPM1-GFP and nuclear marker-RFP were co-expressed in the N. benthamiana leaves, the green and red fluorescent protein signals were superimposed (Fig. 3). Besides, GFP signals were also observed in plasma membrane and cytoplasm under the confocal microscope (Fig. 2d). Above all, CsRPM1 localized in the nucleus, cytoplasm, and cell membrane.
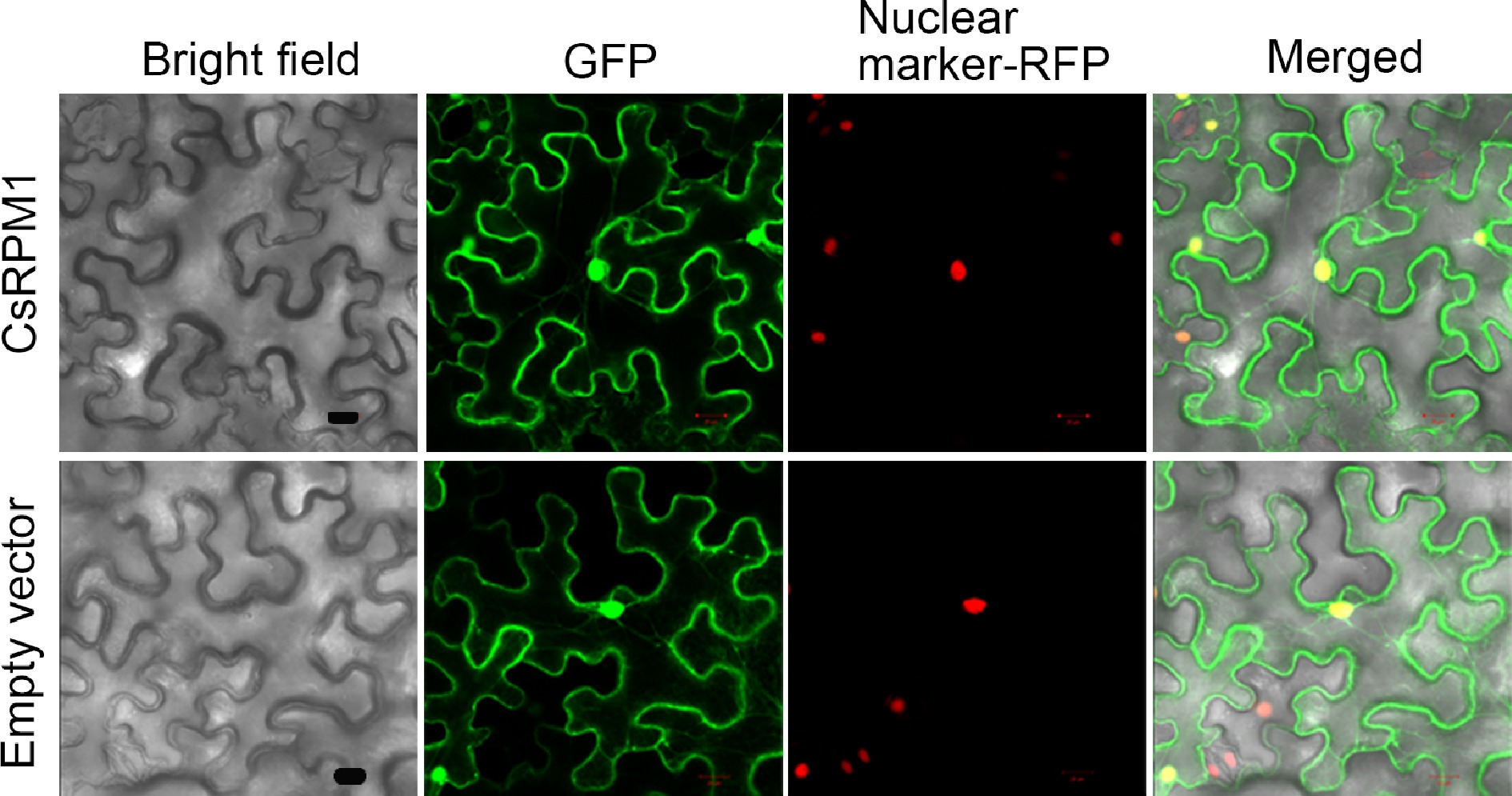
Figure 3.
Subcellular localization of CsRPM1 in the epidermal cells of N. benthamiana leaves. Tobacco leaves expressing nuclear marker-RFP were infiltrated with A. tumefaciens carrying the fused vectors expressing CsRPM1-GFP or empty vector and were observed under laser confocal scanning microscopy after 2 d in the dark. Bar = 50 µm.
MeJA can induce the expression of CsRPM1
-
When analyzing the cis-regulatory elements presenting in the 2 kb upstream region of CsRPM1 that was predicted from LJ43 genomic sequences, we found that MeJA-responsive elements, and defense and stress-responsive elements are presented in the promoter of CsRPM1 gene (Fig. 4a). The transcript level of CsRPM1 was then determined after exogenous treatment with MeJA. qRT-PCR analysis showed that the level of CsRPM1 transcripts was significantly up-regulated at 48 h after MeJA treatment compared with the control group (Fig. 4b). The results indicated that CsRPM1 might be involved in the jasmonic acid (JA) signaling pathway.
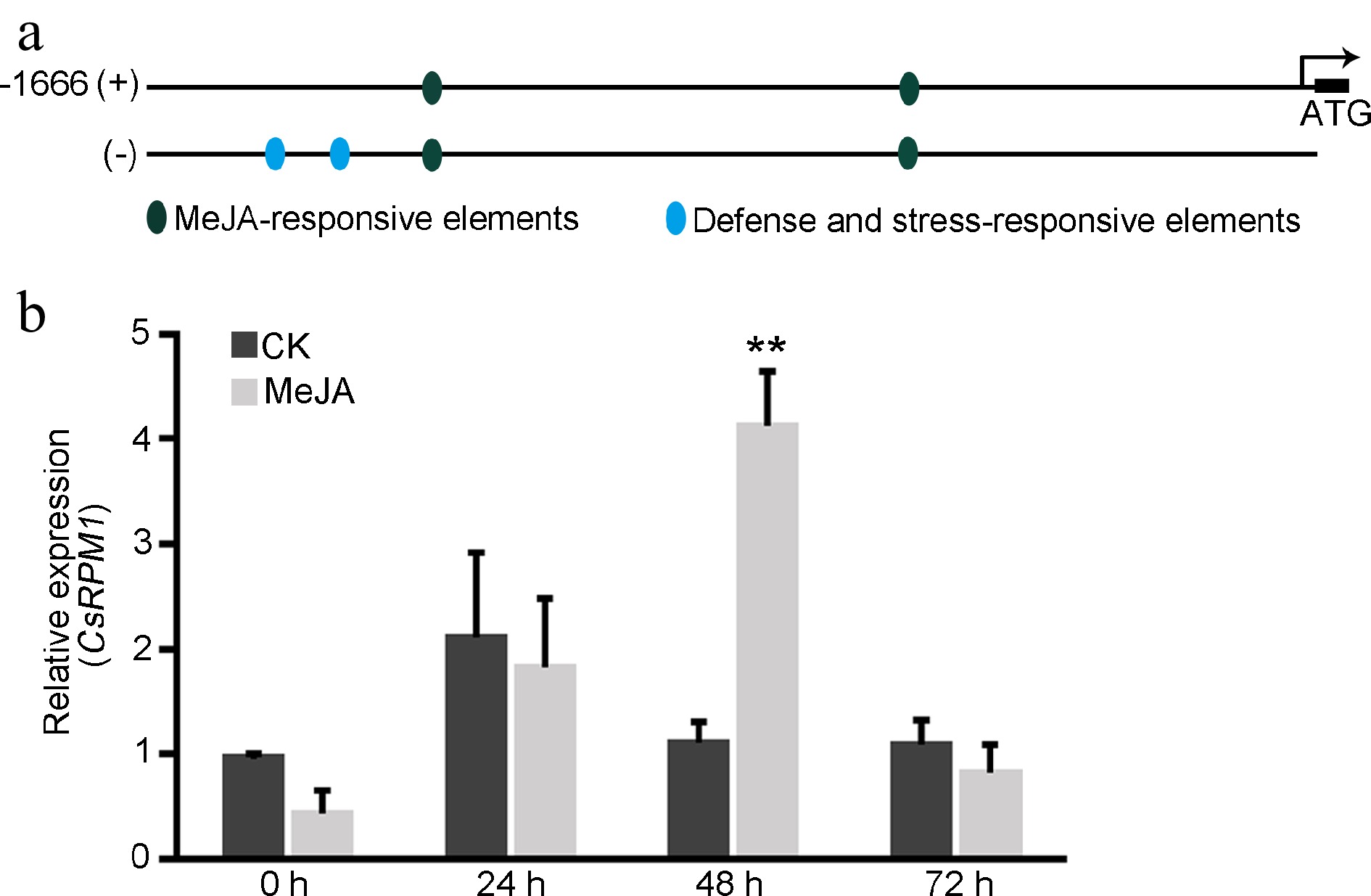
Figure 4.
MeJA can induce the expression of CsRPM1. (a) The predicted promoter elements of CsRPM1 gene from the 2 kb upstream region to ATG. (b) qRT-PCR analysis of CsRPM1 transcripts in tea leaves treated with exogenous 150 μM MeJA. Error bars represent standard deviation. **, p < 0.01.
Predication of candidate transcriptional factors regulating the CsRPM1 expression
-
To further gain insight into the regulatory mechanism of CsRPM1, we predicted the transcriptional factors (TFs) regulating the CsRPM1 expression through the TeaCoN website. A total of 17 DEGs annotated as involved in transcriptional regulation, including 15 up-regulated genes and two down-regulated genes, were identified (Fig. 5a, b). Especially, five genes, TEA030980 (encoding a late embryogenesis abundant protein), TEA031553 (encoding a multicopper oxidase), TEA031563 (encoding a multicopper oxidase), TEA013693 (encoding a calmodulin-binding family protein), and TEA018280 (encoding a VQ motif-containing protein), showed higher correlation with the expression of CsRPM1 and were also significantly up-regulated at 72 h after inoculation with C. camelliae (Fig. 5b). The result suggested that they may be involved in the regulation of CsRPM1 expression.
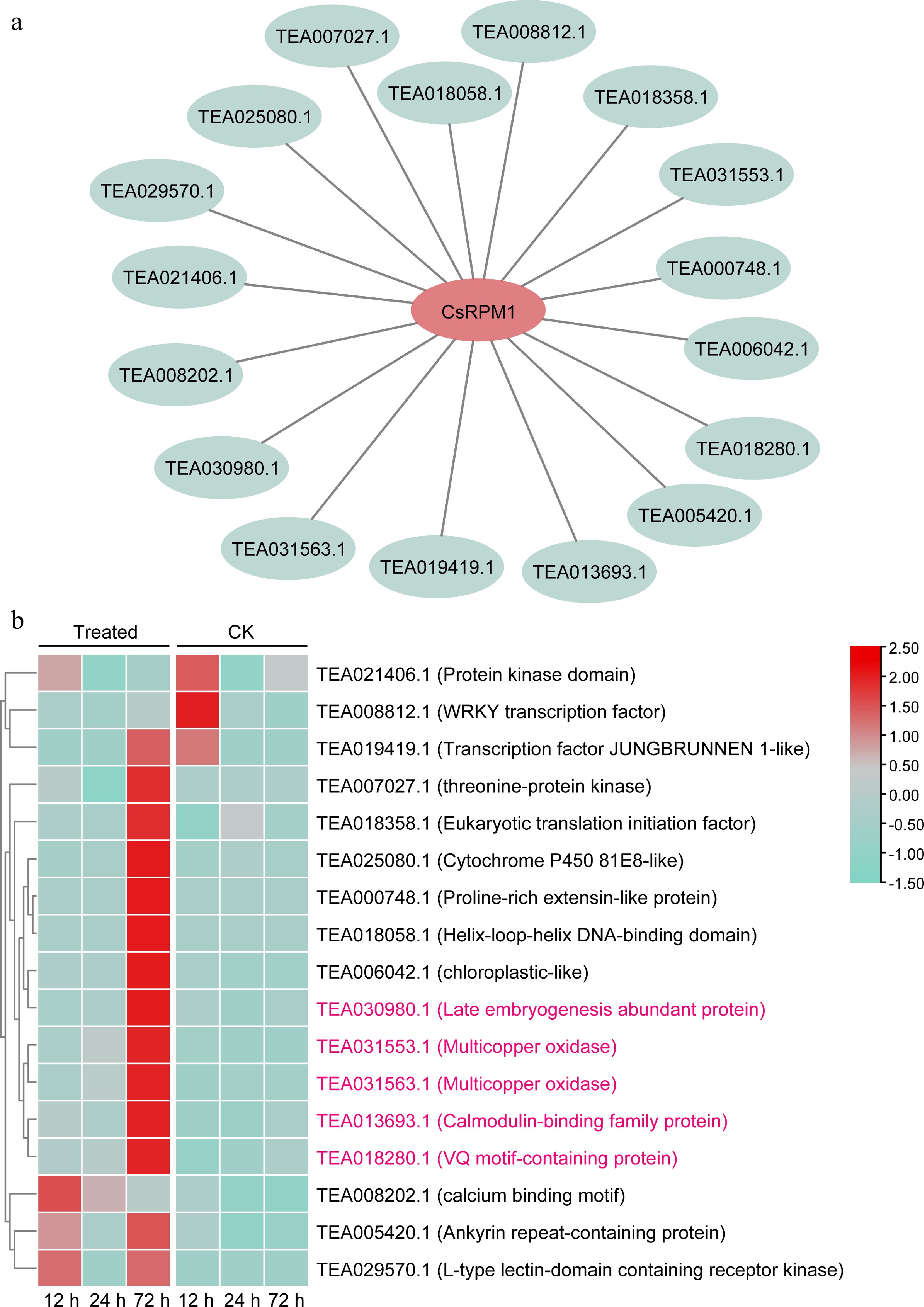
Figure 5.
Connection network between CsRPM1 and candidate TFs. (a) Predicted interaction relationship between CsRPM1 and TFs. (b) Heat map of the expression of candidate TFs in LJ43 at 12 h, 24 h, and 72 h after inoculation with ddH2O (Control) or C. camelliae (Treated).
-
As the most important gene families involved in disease resistance in plants, NLR genes are usually highly expressed following pathogens infection, and the transcript levels of NLRs are correlated with plant defense response against pathogens[9, 13, 25]. In apple, a TIR-NBS-LRR gene MdTNL1 positively regulating Glomerella leaf spot (GLS) resistance was highly expressed in resistance apple cultivar 'Fuji' after inoculation with the GLS pathogen C. fructicola[26]. Powdery mildew resistance gene Pm2b in wheat encoding a CC-NBS-LRR protein was rapidly up-regulated after inoculating with Blumeria graminis f. sp. tritici[27]. Multiple StTNL genes encoding TIR-NBS-LRR proteins in potato showed strong and constant upregulation under fungal pathogen Alternaria solani treatment[28]. In tea plant, RNA-Seq have revealed that the expression of CsNLRs can be induced by anthracnose pathogens[18]. In this study, we identified 12 DEGs commonly expressed at 12 h, 24 h, and 72 h after inoculation with C. camelliae causing anthracnose by RNA-Seq analysis (Fig. 1a), suggesting their potential functions in disease resistance.
Among the 12 DEG
s, CsRPM1 (TEA004876) encoding an NBS-LRR protein was predicted as the disease resistance protein RPM1. RPM1 is an NBS-LRR protein from Arabidopsis that confers resistance to bacterial pathogen Pseudomonas syringae expressing either two avirulence genes, avrRpm1 or avrB[29]. The functional and potentially molecular homologs of RPM1 also exists in conceivably other crop species[30, 31]. In wheat, TaRPM1 plays an important role in the resistance to infection by the powdery mildew pathogen B. graminis f. sp. tritici[32]. PsoRPM1 from Xinjiang wild cherry plum (Prunus sogdiana) is a root-knot nematode resistance gene candidate[33]. In rice, RPM1-like resistance gene 1 OsRLR1 mediates the defense response to the fungal pathogen M. oryzae and the bacterial pathogen Xanthomonas oryzae pv. oryzae through direction interaction with the transcription factor OsWRKY19[17]. In this study, the transcript level of CsRPM1 was significantly upregulated after inoculation with C. camelliae (Fig. 1e), and silencing of CsRPM1 facilitates the infection by C. camelliae and impairs the resistance of tea plant to C. camelliae (Fig. 2a−c). The results indicated that CsRPM1 plays an important role in the resistance to the infection by C. camelliae. RPM1 interacts with other proteins, such as RIN4 and RIN13, which can regulate the RPM1-mediated disease resistance[34, 35]. Therefore, based on the characterization of CsRPM1 function, we can identify proteins directly interacting with CsRPM1 in a yeast two-hybrid screen. Seventeen DEGs including 15 up-regulated genes and two down-regulated genes were predicted to be involved in transcription regulation of CsRPM1 (Fig. 5a, b). Especially, five genes (TEA030980, TEA031553, TEA031563, TEA013693, and TEA018280) were also significantly up-regulated at 72 h after inoculation with C. camelliae (Fig. 5b), showing similar expression patterns to CsRPM1. The results suggested that these genes may be involved in the regulation of CsRPM1 expression. TEA030980 was predicted to encode a late embryogenesis abundant (LEA) protein. In tea plant, LEA gene family are identified and involved in response to abiotic stresses including drought, dehydration, osmotic, and cold[36−39]. However, overexpressing of wheat TdLEA3 in Arabidopsis showed increased resistance to fungal infections by Fusarium graminearum, Botrytis cinerea and Aspergillus niger[40], suggesting its potential role in disease resistance. Multicopper oxidase encoded by TEA031553 or TEA031563 are ubiquitous enzymes that catalyze the oxidation of various substrates by reducing O2 to H2O, including laccases and several oxidases[41]. Calmodulin-binding protein encoded by TEA013693 may act as TF to regulate biotic and abiotic stress responses, especially in low temperature responses[42]. TEA018280 was predicted to encode a VQ (FxxhVQxhTG, h: hydrophobic amino acid, x: any amino acid) motif-containing protein, this family genes in tea plant are identified and found to play important roles in response to salt and drought stress[43]. Although all the five gene families are identified in tea plant, and transcriptome analysis revealed their expression pattern in response to abiotic stresses, their roles in disease resistance to Colletotrichum infection are little known. In future research, we will verify the interaction between CsRPM1 and the five genes by yeast one-hybrid, dual luciferase or electrophoretic mobility shift assays to reveal the regulatory mechanism of CsRPM1 and their roles in disease resistance.
JAs are important phytohormones that regulate a wide range of physiological processes in plant growth and development, and especially the mediation of responses to various stresses[44]. After plants are infected by pathogens, JA and other signaling pathways are activated, which leads to the activation of plant disease resistance[45]. For example, JA biosynthesis and signaling genes in rice can be effectively induced by the rice blast fungus M. oryzae at the warm temperature, which leads to enhanced blast resistance[46]. The JA signaling pathway is involved in PTI and ETI activation, thereby stimulating downstream transcription factors and initiating plant defense responses to pathogen infections[45]. NLRs as the key pathogen effectors activate ETI, many studies have shown that phytohormone pathways including JA signaling pathway are correlated with the resistance mediated by NLRs. In rice, Gene Ontology (GO) analysis revealed strong correlation of hormone pathways, especially salicylic acid (SA) and JA, with the resistance to brown planthopper (BPH) mediated by an NLR gene BPH9[47]. JA and its conjugate JA-Ile rapidly increased after BPH infestation[47]. In tea plant, JA content was also significantly increased in the diseased leaves infected by C. camelliae at 72 h post inoculation[19]. In this study, we have confirmed that the level of CsRPM1 transcripts was significantly up-regulated at 48 h after MeJA treatment (Fig. 4b). These results imply that CsRPM1 may combine with JA signaling pathway to regulate the disease resistance to C. camelliae in tea plant. In cucumber, the transcript levels of two NBS-LRR genes CsRSF1 and CsRSF2 were also upregulated after exogenous treatment with MeJA and correlated with plant defense response against powdery mildew caused by Sphaerotheca fuliginea[9]. The pepper NLR Tsw can recognize pathogen interference of phytohormone signaling as a counter-virulence strategy, thereby activating plant immunity[48]. Therefore, the underlying mechanism of CsRPM1 regulating the defense response to C. camelliae in combination with JA signaling pathway needs to be further explored.
-
NBS-LRR proteins play important roles in plant disease resistance. An NBS-LRR-encoding gene CsRPM1 was identified from tea plant cultivar LJ43. RNA-Seq and qRT-PCR analysis showed that the transcript level of CsRPM1 was up-regulated after inoculation with C. camelliae LS_19 strain. The transient silencing of CsRPM1 in LJ43 leaves comprised the resistance to C. camelliae. The subcellular localization showed that CsRPM1 protein was localized in the nucleus, cytoplasm, and cell membrane. The expression of CsRPM1 was induced by exogenous MeJA. We also predicated the candidate TFs regulating the CsRPM1 expression. Our data indicates that CsRPM1 is required for disease resistance to C. camelliae in tea plant. Further research on the mechanism of CsRPM1 regulating the defense response to C. camelliae in combination with JA signaling pathway and TFs will broaden our comprehension of NLRs in tea plant.
The research was financially supported by the Fundamental Research Funds for the Provincial Universities of Zhejiang (2020YQ001); the Open Fund of Key Laboratory of Biology, Genetics and Breeding of Special Economic Animals and Plants, Ministry of Agriculture and Rural Affairs, P. R. China (TZDZW202202); the Zhejiang Provincial Natural Science Foundation (LY22C160001); Zhejiang Science and Technology Major Program on Agricultural New Variety Breeding-Tea Plant (2021C02067-7).
-
The authors declare that they have no conflict of interest. Xinchao Wang is the Editorial Board member of Beverage Plant Research who was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and his research groups.
-
# These authors contributed equally: Wuyun Lv, Yingjie Xu
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Lv W, Xu Y, Jiang H, Cao Q, Wang X, et al. 2023. An NBS-LRR-encoding gene CsRPM1 confers resistance to the fungus Colletotrichum camelliae in tea plant. Beverage Plant Research 3:13 doi: 10.48130/BPR-2023-0013 

An NBS-LRR-encoding gene CsRPM1 confers resistance to the fungus Colletotrichum camelliae in tea plant
- Received: 15 March 2023
- Revised: 27 April 2023
- Accepted: 09 May 2023
- Published online: 02 June 2023
Abstract: Nucleotide binding site, leucine-rich repeat (NBS-LRR) proteins are the main types of disease resistance proteins in plants, which can perceive plant pathogens. Anthracnose, caused by the fungus Colletotrichum camelliae, is one of the most severe diseases in tea plant. Here, we identified an NBS-LRR-encoding gene, CsRPM1, probably conferring resistance of tea plant to C. camelliae. Phylogenetic analysis showed that CsRPM1 was clustered with RPM1 in Arabidopsis and grouped into CC-NBS-LRR (CNL). It contained a signal peptide, a NB-ARC domain, and multiple LRR motifs. RNA-seq and qRT-PCR analysis showed that the transcript level of CsRPM1 was significantly up-regulated after inoculation with C. camelliae. Transiently silencing of CsRPM1 in tea leaves comprised the resistance to C. camelliae, indicating that CsRPM1 was required for plant defense against the pathogen. The subcellular localization showed that CsRPM1 protein was localized in the nucleus, cytoplasm, and cell membrane. Furthermore, the promoter region of CsRPM1 gene contained MeJA-responsive elements, and the expression of CsRPM1 was induced by exogenous methyl jasmonate, suggesting that CsRPM1 gene may be closely related to JA signaling pathway. A total of 17 transcription factors might be responsible for the expression of CsRPM1. Our data indicates that CsRPM1 is required for disease resistance to C. camelliae in tea plant. The characterization of this disease resistance gene sheds light on NLR protein function in tea plant and may facilitate breeding to control the severe anthracnose.
-
Key words:
- NBS-LRR /
- CsRPM1 /
- Defense /
- Colletotrichum camelliae /
- Tea plant